6. Les protections du système nerveux 6.1 Cellules sensorielles – organes sensoriels

61
6. Les protections du système nerveux
6.1 Cellules sensorielles – organes sensoriels
L’air inspiré subit des turbulences dans la cavité nasale, qui servent à le réchauffer, à l’humidifier,
mais aussi à améliorer les contacts avec l’épithélium olfactif. Cette couche de cellules dans le plafond
de la cavité nasale est recouverte d’une fine pellicule de mucus dans lequel se dissolvent les molécules
volatiles présentes dans l’air.
Des neurones spécialisés de l’épithélium olfactif, les récepteurs olfactifs, portent de fins
prolongements à l’extrémité de leurs dendrites, appelés cils, qui baignent dans le mucus. Lorsque des
molécules odorantes dissoutes dans le mucus entrent en contact avec les cils, les récepteurs olfactifs
concernés sont activés : si le stimulus est suffisant, des potentiels d’actions se forment à leur zone
gâchette. Ces potentiels d’action sont transmis par le nerf olfactif au cerveau qu’ils informent sur la
présence des molécules odorantes perçues.
Les cellules qui, comme les récepteurs olfactifs, réagissent à un stimulus externe par un changement
de leur potentiel de membrane et, finalement, par la formation de potentiels d’action sont appelées
cellules sensorielles.
Figure 51 : Position et structure de l’épithélium olfactif
Chaque cellule sensorielle est activée par une forme particulière de stimulus, son stimulus adéquat.
- Les récepteurs chimiques ou chimiorécepteurs réagissent à des molécules,
- Les photorécepteurs à la lumière,
- Les mécanorécepteurs à des déformations mécaniques,
- Les thermorécepteurs au chaud et au froid.

62
Des cellules sensorielles de même type peuvent constituer des structures complexes avec du tissu non
neuronal : les organes sensoriels.
L’épithélium olfactif constitue un tel organe. Il contient des cellules de soutien qui entourent les
récepteurs olfactifs ainsi que des cellules basales qui, par division cellulaire, forment de nouveaux
récepteurs olfactifs, permettant ainsi leur continuel remplacement.
Quel que soient le stimulus ou les processus se déroulant dans les cellules sensorielles, le principe de
base reste toujours le même : la cellule sensorielle répond à une stimulation par un changement de son
potentiel de membrane. Il se forme d’abord un potentiel de récepteur dont l’amplitude est
proportionnelle à l’intensité du stimulus. Ce processus – la transformation d’un stimulus en un
potentiel de récepteur – caractérise les cellules sensorielles et est appelé transduction. Lorsque
l’intensité du stimulus dépasse une certaine valeur, appelée seuil de stimulation, le potentiel de
récepteur devient suffisant pour générer des potentiels d’action qui sont conduits au système nerveux
central. La fréquence des potentiels d’action code l’intensité du stimulus.
Figure 52 : Transduction dans une cellule olfactive. A Potentiel de récepteur et potentiels d’action –
B Cascade de signaux dans la membrane des cil.
La seule information transmise par les cellules sensorielles au cerveau concerne
donc l’intensité du stimulus. Pourtant, les potentiels d’action parvenant au
cerveau y induisent des perceptions sensorielles fort diversifiées.
Cela résulte du fait que les potentiels d’action atteignent le cerveau par
différentes voies et qu’ils y activent différentes régions spécifiques.
On distingue deux types de cellules sensorielles : celles qui, comme les
récepteurs olfactifs, produisent des potentiels d’action qui parviennent
directement au cerveau sont appelées cellules sensorielles primaires. Ce sont en
fait des neurones transformés. Les autres sont des cellules épithéliales
modifiées ; on les appelle cellules sensorielles secondaires. Dans leur cas, le
potentiel de récepteur conduit à la libération de neurotransmetteurs qui agissent
sur un neurone. C’est celui-ci qui produit les potentiels d’action.
La transduction se déroule différemment selon le type de récepteur sensoriel.
Toutefois, des canaux ioniques spécialisés sont toujours impliqués. Ils sont
activés soit directement par le stimulus, soit par une chaîne de signaux
cellulaires. Une telle chaîne est impliquée dans les récepteurs olfactifs lorsqu’une Figure 53 : Type
de cellules
sensorielles

63
molécule odorante s’attache à une protéine réceptrice, présente dans la membrane des cils, et
spécifique de la molécule en question. Chaque récepteur olfactif ne contient qu’un seul type de
protéine réceptrice de l’odeur, qui présente une affinité différente à diverses molécules odorantes. Si
une protéine réceptrice de l’odeur s’attache à une molécule odorante, elle active alors une protéine G
qui elle-même active une adénylate cyclase. Cette enzyme membranaire catalyse la formation d’AMP
cyclique (AMPc) à partir d’ATP. L’AMPc agit comme un second messager. Il permet la transmission
à l’intérieur de la cellule sensorielle de signaux provenant de stimuli- les substances odorantes – qui
restent à l’extérieur de celle-ci. La présence d’AMPc entraîne l’ouverture de canaux cationiques
spécifiques, ce qui génère un courant d’ions sodium, et surtout calcium, vers l’intérieur du cil. Un
potentiel de récepteur est produit. L’augmentation d’ions calcium dans les cils conduit à l’ouverture de
canaux chlorure. Le courant d’ions chlorure vers l’extérieur de la cellule renforce le potentiel de
récepteur et aboutit à la formation de potentiels d’action au niveau de la zone gâchette. Ces potentiels
d’action sont ensuite transmis au cerveau. Comme la liaison d’une seule molécule odorante à une
protéine réceptrice de l’odeur conduit à la production de nombreuses molécules d’AMPc et comme
chacune de ces molécules ouvre plus d’un canal ionique, la chaîne de signaux implique une importante
amplification du signal ; c’est pourquoi on parle d’une cascade de signaux.
Les récepteurs olfactifs peuvent s’adapter, c’est-à-dire qu’ils peuvent changer leur sensibilité à un
stimulus. Lors d’une stimulation continue par une substance odorante, la cellule ne produit des
potentiels d’action que durant une seconde environ. Il faut ensuite que l’intensité du stimulus change
pour obtenir de nouvelles réponses.
6.2 L’œil – un organe photosensible
L’œil humain (figure 54) – comme celui de tous les vertébrés – est un organe photosensible complexe,
constitué de photorécepteurs, de cellules nerveuses et de structures transparentes réfractant la lumière :
- la cornée,
- l’humeur aqueuse,
- la pupille,
- le cristallin,
- le corps vitré.
Figure 54 : L’œil humain. A Coupe horizontale de l’œil gauche – B Accommodation. A gauche : vue
frontale, à droite : coupe sagittale.
Ensemble, ces structures forment le système optique de l’œil ou appareil dioptrique, qui projette sur
la rétine une image réduite et inversée des objets placés devant l’œil. Le cristallin peut se déformer, ce

64
qui change son pouvoir de réfraction ou convergence. C’est ce qui permet d’obtenir des images nettes
d’objets placés à différentes distances de l’œil. Pour la vision lointaine, le cristallin est aplati, pour la
vision proche, il est fortement bombé. Cette capacité du cristallin à changer de forme résulte de
l’action combinée des muscles ciliaires, de la choroïde et des ligaments suspenseurs du cristallin.
L’image d’un objet éloigné est nette pour un œil au repos. Les muscles ciliaires sont relâchés et les
ligaments suspenseurs du cristallin sont tendus par la traction de la choroïde : le cristallin est aplati et
son pouvoir de réfraction est faible. Si l’image des objets éloignés est nette sur la rétine, celle des
objets proches est floue. Si l’on fixe un objet proche de l’œil, les muscles ciliaires, qui encerclent le
cristallin, se contractent et diminuent la traction que les ligaments suspenseurs exercent sur le
cristallin. Celui-ci se bombe jusqu’à prendre une forme presque sphérique sous l’effet de son
élasticité. Le pouvoir de réfraction du cristallin augmente et une image nette de l’objet proche se
forme sur la rétine. En revanche, le fond du champ visuel forme une image floue. On appelle
accommodation cette capacité de l’œil à former une image nette sur la rétine d’objets très proches ou
éloignés. Avec l’âge, l’élasticité du cristallin diminue, ce qui empêche la vision nette d’objets
rapprochés ; c’est la presbytie, qui est corrigée par le port de lunettes convergentes.
L’ouverture de la pupille peut aussi être modifiée. La pupille se réduit par suite de la contraction des
fibres musculaires circulaires de l’iris, alors qu’elle s’élargit par la contraction des fibres radiales. Par
ce mécanisme, l’œil s’adapte aux changements de luminosité. L’ouverture de la pupille se réduit aussi
lors de l’accommodation à une vision rapprochée, ce qui augmente la profondeur de champ et donc la
netteté de l’image.
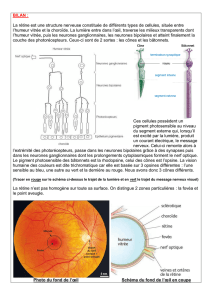
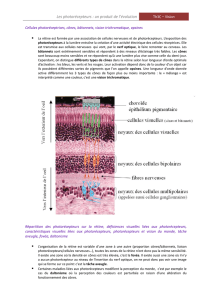
La rétine (figure 28) contient des
photorécepteurs et des neurones
qui captent l’image qui y est
projetée par l’appareil dioptrique
et la transmettent au cerveau sous
forme de potentiels d’action. La
rétine est composée de plusieurs
couches cellulaires. Un épithélium
pigmenté, en contact avec la
choroïde, forme la couche externe.
Le pigment (mélanine) absorbe la
lumière qui diffuse et empêche la
formation de reflets. Les personnes
atteintes d’albinisme n’ont pas de
mélanine et sont éblouies en forte
lumière. La couche suivante, en
direction de l’intérieur de l’œil, est
formée par les
photorécepteurs (figure 56): les
cônes et les bâtonnets. Les cellules
nerveuses viennent ensuite. Les photorécepteurs sont couplés par des synapses aux cellules
bipolaires, qui elles-mêmes sont reliées aux cellules ganglionnaires dont les axones forment le nerf
optique. Les cellules horizontales établissent des contacts latéraux entre les photorécepteurs et les
cellules amacrines entre les cellules bipolaires.
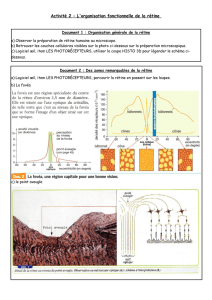
Figure 56 : Structure des photorécepteurs chez les vertébrés. A Bâtonnet – B Cône
Figure 55 : Structure de la rétine chez les vertébrés

65
Les photorécepteurs sont
absents de l’endroit où le
nerf optique quitte l’œil
(figure 57) : c’est la
tache aveugle. L’endroit
où l’acuité visuelle est
maximale est appelée la
tache jaune. Les
cellules nerveuses qui,
ailleurs sur la rétine, sont
placées devant les
photorécepteurs sont ici
placées sur leur côté, ce
qui permet à la lumière
d’atteindre directement
les photorécepteurs. La
position des cellules
nerveuses de la tache
jaune conduit à la
formation d’une
dépression : la fovéa
centralis (lat. fovea :
trou), abrégée fovéa. Les
photorécepteurs
présentes au niveau de la
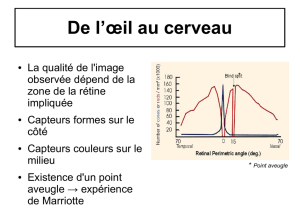
tache jaune ne sont que des cônes ; plus on s’en éloigne, plus la proportion de bâtonnets augmente. Le
rapport entre le nombre de photorécepteurs et le nombre de cellules ganglionnaires varie aussi : alors
qu’au niveau de la fovéa chaque photorécepteurs couplé à sa propre cellule ganglionnaire, en
périphérie, plusieurs photorécepteurs sont couplés à une même cellule ganglionnaire.
Les photorécepteurs sont divisés en deux segments (figure 56). Dans le segment interne se trouvent le
noyau, les mitochondries et le réticulum endoplasmique. Ce segment se termine par une structure
présynaptique, où se fait le contact avec les neurones bipolaires. Le segment externe est rempli de
disques membranaires qui contiennent les pigments photosensibles. Les bâtonnets et les cônes se
distinguent par la forme et la structure de leur segment externe. Un bâtonnet possède un segment
externe long, cylindrique et contenant de nombreux disques. Le segment externe d’un cône est plus
court, conique, contient moins de disques et donc moins de pigments photosensibles. En conséquence,
les cônes sont moins sensibles que les bâtonnets. Par contre, si la luminosité est suffisante, ce sont eux
qui transmettent l’information concernant les couleurs. Les bâtonnets ont la capacité de former une
image encore assez nette en faible lumière, mais seulement dans des nuances de gris.
6.3 La phototransduction
Alors que le potentiel de repos des neurones est de -70mV, le potentiel de repos d’un bâtonnet à
l’obscurité avoisine -30mV (figure 58a). Cela s’explique par une entrée continue de sodium par des
canaux sodium particuliers, placés dans la membrane du segment externe. Ces canaux sont maintenus
ouverts en présence de guanosine monophosphate cyclique (GMPc), un messager secondaire. A cette
valeur du potentiel de membrane des bâtonnets, les synapses reliant ces derniers avec leurs cellules
bipolaires libèrent continuellement le neurotransmetteur glutamate. Or, les canaux sodium présents
dans la membrane postsynaptique sont maintenus fermés en présence de glutamate : en conséquence,
la cellule bipolaire reste inactive.
Figure 57 : Distribution des cônes et des bâtonnets sur la rétine
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%