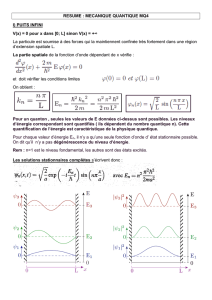
Contamination du marsouin commun Phocoena phocoena des perturbateurs endocriniens persistants

FACULTE DE MEDECINE VETERINAIRE
Département des Sciences des Denrées alimentaires
Section d’Analyse des Denrées alimentaires
Contamination du marsouin commun
(Phocoena phocoena) de la Mer du Nord par
des perturbateurs endocriniens persistants
Persistent endocrine disruptors contamination
in North Sea harbour porpoises
(Phocoena phocoena)
Pedro Henrique DIDIMO IMAZAKI
Mémoire présenté en vue de l’obtention
du Diplôme de Master Complémentaire en Médecine vétérinaire spécialisée
Option Santé publique vétérinaire : Sciences des Aliments
Année académique 2008 – 2009

Contamination du marsouin commun (Phocoena
phocoena) de la Mer du Nord par des
perturbateurs endocriniens persistants
Mots-clés : perturbateurs endocriniens, marsouin commun, Mer du Nord, composés
organochlorés, chromatographie en phase gazeuse, spectrométrie de masse, gène
rapporteur
Persistent endocrine disruptors contamination in
North Sea harbour porpoises
(Phocoena phocoena)
Keywords: endocrine disrupters, harbour porpoise, North Sea, organochlorine
compounds, gas chromatography, mass spectrometry, reporter gene

Le Professeur Marie-Louise Scippo, promotrice de ce mémoire présenté en vue de
l’obtention du diplôme de Master complémentaire en Médecine vétérinaire
spécialisée / Option Santé Publique vétérinaire : Sciences des Aliments, a accepté la
publication de cet ouvrage. L’auteur et le promoteur autorisent sa consultation et sa
photocopie pour usage personnel.
Toute utilisation doit se conformer aux dispositions sur les droits d’auteurs, en
particulier l’obligation de mentionner la source lors de l’utilisation de résultats de
cette étude.

REMERCIEMENTS
Je voudrais tout d’abord remercier le Prof. Marie-Louise Scippo pour m’avoir
accueilli dans son laboratoire, pour m’avoir donné l’occasion de réaliser ce mémoire
et surtout pour sa patience et pour sa disponibilité.
Je voudrais ensuite remercier tout le personnel du Laboratoire d’analyses de
denrées alimentaires pour leur encouragement, pour leur amitié et pour ces deux
années de moments inoubliables.
Finalement, un grand merci à ma famille et à mes amis, qui m’ont soutenu
tout au long de la réalisation de ce travail.

AVANT-PROPOS
« (…) on peut produire autrement, durablement,
des denrées alimentaires de haute qualité
sanitaire, en quantité suffisante, avec des
performances économiques bien supérieures
aux méthodes industrielles, pour peu que l’on
cesse de remplacer les paysans sur la Terre
par des molécules chimiques. »
(Philippe Desbrosses)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
1
/
100
100%