Neurologie [17-013-A-10] Troubles de la sensibilité Henri Dehen

Neurologie
[17-013-A-10]
Troubles de la sensibilité
Henri Dehen : Professeur de neurologie à la faculté Xavier-Bichat (16, rue Henri-
Huchard, 75018 Paris)
Clinique neurologique, hôpital Beaujon, 100, boulevard du Général Leclerc, 92110
Clichy France
Thomas De Broucker : Praticien hospitalier, chef de service
Service de neurologie, hôpital Delafontaine, 93205 Saint-Denis cedex 1 France
Résumé
De nombreux travaux ont, au cours de ces 20 dernières années, modifié profondément
nos connaissances sur la sensibilité et ses troubles. De nouvelles techniques
neuroanatomiques, électrophysiologiques et neurochimiques ont notamment permis de
mieux comprendre l'élaboration et la transmission des messages somesthésiques.
Ainsi, l'enregistrement des potentiels évoqués somesthésiques chez l'homme est
devenu une pratique courante. Par ailleurs, et peut-être surtout, une meilleure
connaissance des systèmes de contrôle physiologique de la sensation douloureuse a
conduit à de nouvelles stratégies thérapeutiques et à une modification du
comportement médical vis-à-vis de la douleur rebelle.
Organisation anatomofonctionnelle des systèmes sensitifs [5
Bernard JF, Besson JM The spino (trigemino)-pontoamygdaloid pathway : electrophysiological evidence for an involvement in pain processes. J
Neurophysiol 1990 ; 63 : 473-490

Cliquez ici pour aller à la section Références, 8
Besson JM, Chaouch A Peripheral and spinal mechanisms of nociception. Physiol Rev 1987 ; 67 : 67-186
Cliquez ici pour aller à la section Références, 11
Cervero F, Iggo A The substantia gelatinosa of the spinal cord - a critical review. Brain 1980 ; 103 : 717-772
Cliquez ici pour aller à la section Références, 30
Iggo A. Somatosensory system. In : Handbook of sensory physiology. New York, Springer Verlag : 1973
Cliquez ici pour aller à la section Références, 32
Jones AK, Brown WD, Friston KJ, Qi LY Frackowiak RS. Cortical and subcortical localization of response to pain in man using positron emission
tomography Proc R Soc Lond Biol 1991 ; 244 : 39-44 [crossref]
Cliquez ici pour aller à la section Références, 48
Talbot JD, Marrett S, Evans AC, Meyer E, Bushnell MC, Duncan GH Multiple representations of pain in human cerebral cortex. Science 1991 ; 251 :
1355-1358
Cliquez ici pour aller à la section Références, 55]
Willis WD, Coggeshall RE. Sensory mechanisms of the spinal cord (2nd ed). New York, London, Plenum Press : 1991 : 575 p
Cliquez ici pour aller à la section Références
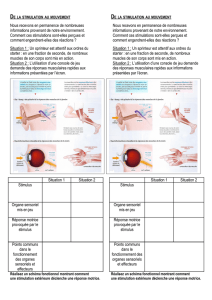
Système périphérique de réception
Transduction

Elle est le premier pas de l'information sensorielle. Il s'agit de la conversion en activité
neuronale de l'énergie naturelle transmise au récepteur. L'activation naturelle, pour être
convenablement transmise, doit être d'intensité suffisante (supérieure au seuil) et de la
qualité voulue en rapport avec le type de récepteur. Le champ récepteur excède celui du
tissu directement innervé car les tissus cutanés avoisinants peuvent transmettre l'énergie
du stimulus. La transduction sensorielle va aboutir au potentiel de récepteur dont le rôle
dans l'initiation de l'influx nerveux est comparable à celui du potentiel postsynaptique
d'excitation. Le potentiel de récepteur est produit par une modification des perméabilités
membranaires au Na+ et au K+. Le potentiel de récepteur est un phénomène local, non
propagé, le déclenchement d'un potentiel d'action, propagé, dépend du dépassement du
seuil par le potentiel de récepteur.
Lorsque la stimulation est supraliminaire, le potentiel de récepteur croît plus rapidement et
a une amplitude plus grande, évoquant alors des trains de potentiels d'action dont la
fréquence croît avec l'intensité de la stimulation.
Codage
Celui de l'information sensitive se fait d'une part par la fréquence de décharge de chaque
neurone spécialisé dans le mode de stimulation, d'autre part par le recrutement d'une
population croissante de neurones.
Une certaine forme de codage est liée aux phénomènes d'adaptation des récepteurs.
Tous les récepteurs s'adaptent lorsque la stimulation est prolongée. Mais cette adaptation,
qui se traduit par une diminution du potentiel de récepteur et donc par une diminution de la
fréquence des potentiels d'action, est de rapidité variable en fonction du type de récepteur.
Un type de récepteur d'adaptation lente est le fuseau neuromusculaire. Un type de
récepteur d'adaptation rapide est le corpuscule de Pacini ; dans ce cas, l'enveloppe
conjonctive de sa terminaison nerveuse est à l'origine des caractéristiques d'adaptation :
la décharge neuronale liée à l'activation mécanique ne surviendra qu'au début et parfois à
la fin du stimulus (indentation cutanée).
Activité neuronale
Évoquée par les stimulations périphériques, elle est véhiculée par des fibres de différents
diamètres, conduisant donc à des vitesses différentes.
Les histogrammes des nerfs afférents : nerfs musculaires et nerfs cutanés, sont
différents ; les nerfs afférents musculaires contiennent quatre types de fibres :
I : myélinisées de grand diamètre ;
II : myélinisées de petit diamètre ;

III : myélinisées de plus petit diamètre ;
IV : amyéliniques. La nomenclature utilisée pour les afférences sensitives cutanées est
différente (Aα β δ et C), les fibres Aα sont absentes de ces afférences cutanées.
Des travaux électrophysiologiques récents, en montrant la spécificité de la sensation en
cas d'activation d'un récepteur donné ou de sa fibre afférente, plaident en faveur de la
théorie de la spécificité qualitative de la sensation qui paraît donc rendre compte de la
grande majorité des processus de codage des stimuli sensitifs.
Différentes catégories de récepteurs
Nocicepteurs
Ils ont comme caractéristique d'être activés par des stimulations pour lesquelles le seuil
d'excitation est plus haut que pour les autres catégories de récepteurs. En règle générale,
l'intensité nécessaire correspond à une stimulation nocive, c'est-à-dire potentiellement
lésante.
Leurs fibres afférentes sont de type Aδ (4-36 m/s) et C (0,5-2 m/s). Deux grandes classes
de nocicepteurs sont individualisées : les mécanonocicepteurs (fibres Aδ) et les
nocicepteurs polymodaux (fibres C). Mais cette classification est soumise à controverse du
fait de l'activation de fibres Aδ par certains nocicepteurs polymodaux et de l'existence
possible de certains nocicepteurs sensibles électivement aux stimulations thermiques.
Les mécanonocicepteurs sont activés par des stimulations mécaniques intenses
uniquement (pincement, piqûre) et sont associés à des fibres afférentes Aδ. Celles-ci
représentent 20 % des fibres Aδ d'origine cutanée. Leur champ récepteur est assez large
(1 à 2 cm).
Les nocicepteurs polymodaux sont associés à des fibres du groupe C et répondent à des
stimulations de tous types : mécaniques intenses, thermiques (inférieure à 18 °C ou
supérieure à 42 °C) ou chimiques. Leur champ récepteur est en général de petite
dimension. Leur réponse présente un seuil élevé et croît avec l'intensité de la stimulation.
L'existence de ces deux types de récepteurs rend compte du phénomène de la double
douleur ressentie lors d'un stimulus ponctuel comme un pincement : douleur rapide, aiguë,
localisatrice, suivie d'une douleur retardée ou lente, pénible à type de brûlure, moins
localisable et plus durable. Ce manichéisme doit être tempéré par la possibilité d'activation
de fibres Aδ par les récepteurs polymodaux d'une part, d'autre part par l'existence d'autres
types de récepteurs activés par les stimulations nocives, en particulier thermiques. L'étude
des nocicepteurs d'origine musculaire, articulaire ou viscérale est plus délicate. Il a été mis
en évidence des nocicepteurs polymodaux Aδ et C au niveau des articulations et des
muscles chez l'animal. Mais, en ce qui concerne les récepteurs viscéraux, il n'est pas

encore établi si la douleur résulte de nocicepteurs spécifiques ou s'il s'agit de
l'accroissement de l'activité de récepteurs impliqués dans l'activité physiologique.
Le mécanisme de l'activation des nocicepteurs n'est pas établi avec certitude, toutefois :
l'action de substances algogènes telles que bradykinine, histamine, sérotonine, K+...
dans la décharge des nocicepteurs est clairement démontrée ainsi que le rôle
sensibilisateur des prostaglandines au niveau de la fibre nerveuse ;
l'activation d'un nocicepteur entraîne par réflexe d'axone une vasodilatation de
voisinage avec inflammation et activation de nouveaux nocicepteurs. La substance
P a été incriminée dans ce phénomène.
Thermorécepteurs
Les thermorécepteurs ont une sensibilité élective à la température et aux changements de
température de la peau. Ils sont presque totalement insensibles aux stimulations
mécaniques.
La répartition cutanée des thermorécepteurs est responsable du caractère ponctué de la
thermosensibilité, par " taches " de 1 mm de diamètre environ, dont la stimulation entraîne
une sensation soit de froid, soit de chaud. La sensibilité des récepteurs est maximale entre
23 et 36 °C pour les récepteurs au froid et autour de 40 °C pour les récepteurs au chaud.
En outre, au-delà de 45 °C, les récepteurs au froid répondent de nouveau, de façon
paradoxale, responsables d'une sensation de froid lorsqu'un spot est stimulé par une
chaleur douloureuse focalisée. Les fibres nerveuses innervant ces récepteurs sont de
types Aδ et C pour les récepteurs au froid et C pour ceux au chaud.
Mécanorécepteurs
Ils véhiculent les sensations tactiles et sont innervés par des fibres Aβ à conduction
rapide.
La peau glabre contient quatre types de récepteurs mécaniques :
deux sortes de récepteurs d'adaptation rapide caractérisés
morphologiquement : les corpuscules de Meissner sont situés dans les
papilles dermiques et ont un champ récepteur de petite taille alors que les
corpuscules de Pacini se trouvent dans le tissu sous-cutané et ont un champ
de réception large ;
deux sortes de récepteurs d'adaptation lente distincts eux aussi
morphologiquement : les corpuscules de Merkel et de Ruffini ; ces derniers
ont un champ récepteur plus large que les corpuscules de Merkel.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
1
/
51
100%


![21.Francis PONGE : Le parti pris de choses [1942]](http://s1.studylibfr.com/store/data/005392976_1-266375d5008a3ea35cda53eb933fb5ea-300x300.png)