Contraction musculaire du MSS

République Algérienne et Populaire
Ministère de l’enseignement supérieur et de le recherche
scientifique
Université de Mostaganem
Physiologie de la contraction
musculaire du MSS
Présenté par Dr Benchohra
Année Universitaire 2014-2015

Plan
•I-Introduction.
•II-Rappel sur la transmission neuromusculaire.
•III- Le couplage excitation-contraction : la
contractilité.
•IV-Les sources d énergie de la contraction
musculaire.
•V-Conclusion.

I/-Introduction:
Les cellules musculaires ont 2 propriétés fondamentales :
–L’excitabilité .
–La contractilité.
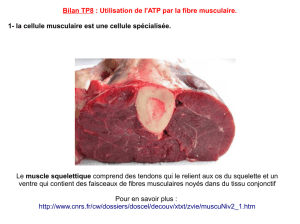
•La cellule musculaire est dite excitable car elle répond
de façon spécifique au potentiel d’action nerveux qui
va générer un potentiel d’action musculaire : on parle
des mécanismes de transmission neuro-musculaire.
•L’apparition du PA mettra en jeu une série de
mécanismes contractiles que l’on décrit sous le terme
de couplage excitation / contraction, qui aboutira à la
création d’une tension musculaire.

II-Rappel sur la transmission neuro
musculaire:

1/-L’innervation motrice :
Les fibres musculaires sont innervées par des fibres
nerveuses myélinisées de gros calibre et ayant
des vitesses de conduction élevées, comprises
entre 60 et 120 m/s. Elles font partie des plus
grosses fibres nerveuses de notre organisme.
•On appelle ces fibres nerveuses des
motoneurones A, dont le corps cellulaire se
situe au niveau de la corne antérieure de la
moelle épinière.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
1
/
31
100%