Blanc, F., Ledeme, P. Blanc, Ch. (1987). Quelques r sultats des travaux men s sur la variabilit g n tique chez la perdrix grise. Bulletin Mensuel de l O.N.C., 113 : 11-13.

r›
v
l
Quelques
résultats
des
travaux
menés
sur
la
variabilité
génétique
chez
la
perdrix
grise
INTRODUCTION
Un
travail
de
recherche
a
été
mené
par
le
Laboratoire
de
Zoo-
géographie
de
l'Université
de
Montpellier,
sur
la
variabilité
géné-
tique
de
la
Perdrix
grise,
à
la
demande
de
l'Office
National
de
la
Chasse.
Les
méthodes
d'analyses
couram-
ment
employées
par
ailleurs
sur
des
espèces
animales
par
des
laboratoires
de
recherche
spécia-
lisés
ont
ainsi
été
appliquées
à
cette
espèce.
La
question
posée
était
de
savoir
si
on
pouvait
mettre
en
évidence
des
particularités
génétiques
chez
les
perdrix
grises
des
Pyrénées
qui
les
distinguent
des
perdrix
grises
de
plaine.
Ce
travail
a
répondu
positivement
à
cette
question.
Il
a
également
montré quels
pouvaient
être
les
apports
et
les
limites
de
travaux
de
ce
type
pour
les
gestionnaires
de
la
faune.
Des
confirmations
devront
être
apportées
en
multipliant
les
échan-
tillons
et
en
recherchant
le
maxi-
mum
de
précisions
biologiques
et
écologiques
sur
les
populations
étudiées.
Photo
O.N.C.
B.M.
O.N.C.
N°113.
Mai
1987
Un
rappel
de
définitions
de
termes
génétiques
employés
dans
ce
texte
est
sans
doute
nécessaire
au
non
spécialiste
pour
comprendre
les
résultats
présentés:
Gènes
Les
gènes
sont
les
supports
des
caracteres
d'un
individu.
Ils
lui
sont
transmis
par
ses
parents.
Par
exemple,
le
caractère
de
couleur
de
la
lleur
«Belle
de
nuit»
est
porté
par
un
gène
qu'on
peut
appeler
le
gène
«couleur
de
la
lleur››.
Locus
Dans
chaque
cellule
d'un
organisme,
les
gènes
sont
portés
par
les
chromosomes.
La
position
qu'ils
occupent
sur
ces
chromosomes
est
appelée
locus;
c'est
le lieu
où
ils
se
trouvent.
Les
chromosomes
étant en
double
dans
chaque
cellule,
chaque
caractere
est
représenté
par
deux
gènes.
Dans
notre
exemple,
le
caractère
de
la
lleur
est
donc
supporté
par
deux
gènes.
Hllèle
Ces
deux
gènes
peuvent
être
identiques
ou
de
type
différent.
On
appelle
allèle,
ces
différents
types
d'un
même
gène.
Les
genes
situés
sur
un
même
locus
sont
des
allèles.
Dans
notre
exemple,
le
gène
«couleur
de
la
lleur»
comporte
deux
allèles
R
et
B.
Génotype
Les
deux
gènes
supportant
un
même
caractère
forment
le
génotype
cle
l'indi-
vidu
pour
ce
caractère.
Dans
notre
exemple,
les
génotypes
seront
de
trois
types:
RR,
RB
ou
BB.
Une
fleur
aura,
pour
le
caractère
couleur,
un
génotype
RR,
RB
ou
BB.
Phénoty-pe
L'expression,
ou
réalisation,
d'un
génotype
est
appelé
phénotype.
Dans
notre
exemple,
les
individus
présenteront
trois
phénotypes
différents:
0
couleur
rouge
(génotype
RR),
0
couleur
rose
(génotype
RB),
0
couleur
blanche
(génotype
BB).
Population
C'est
la
collectivité
la
plus
élémentaire
au
sein
de
laquelle
les
échanges
gené-
tiques
sont
absolument
libres
et
constants
(Dobzhansky,
1952).
Exemple
les
«Belles
de
Nuit»
d'une
même
serre.
Polymorphisme
allélique
Si
cette
population
présente
une
fréquence
élevée
du
taux
de
présence
de
cha-
cune
des
allèles,
ici
R
ou
B,
on
dira
qu'elle
présente
un
taux
de
polymorphisme
allélique
élevé
(par
exemple:
45%
de
R
et
55%
de
B).
A
l'inverse,
si
les
proportions
étaient
de
98%
de
R
et
2%
de
B,
on
aurait
une
population
à
faible
taux
de
polymorphisme
allélique,
puisqu'il
n'y
aurait
prali-
quement
que
l'allèle
R
qui
serait
représenté.
Polymorphisme
génotypique
Si
cette
population
présente
une
fréquence
élevée
du
taux
de
présence
de
cha~
cun des
génotypes,
ici
RR,
RB
ou
BB,
on
dira
qu'elle
présente
un
taux
de
poly-
morphisme
génotypique
élevé
(par
exemple:
RR
=
30%
_
RB
=
35%
-
BB
=
35%).
A
l'inverse,
une
population
présentant
cles
proportions
de
96%
pour
le
génotype
RR,
2%
pour
RB
et
2%
pour
BB,
serait
presque
monomorphe
et
son
taux
de
polymorphisme
génotypique,
faible.
G.
CHANTREL
11

PROBLÉMATIQU1:
La
Perdrix
grise
est
présente
dans
les
plaines
de
toute
la
partie
Nord
de
la
France
et
en
montagne
dans
les
Pyrénées
et
dans
les
Alpes.
On
peut
considérer
qu'au
Nord
d'une
ligne
Bordeaux-Genève,
la
Perdrix
grise
est
présente
partout,
alors
qu'en
altitude
elle
n'existe
que
dans
certains
secteurs
limités.
L'aire
de
répartition
géographique
de
cette
espece
est
donc
disconti-
nue
et,
qui
plus
est,
elle
présente
des
îlots
bien
séparés,
en
particu-
lier
dans
la
chaîne
Pyrénéenne.
Cette
répartition
particulière
peut
correspondre
à
des
compositions
génétiques
différentes
entre
les
diverses
populations
de
perdrix
grises.
On
serait
alors
en
présence
de
«souches»
présentant
chacune
des
caractéristiques
génétiques
particulières.
On
a
cherché
à
mettre
en
évi-
dence
l'existence
de
ces
différen-
ces
dans
la
mesure
ou
si
elles
exis-
tent
vraiment,
elles
devront
être
prises
en
compte
dans
la
gestion
de
l'espece;
il
faudrait
veiller
à
ce
que
des
souches
locales
ne
dispa-
raissent
pas
et
il
serait
souhaitable
d'éviter
l'introduction
d'oiseaux
«étrangers»
au
sein
d'une
souche
et
en
particulier
éviter
le
lâcher
d'oiseaux
d'élevage.
Uétude
qui
a
été
entreprise
se
pro-
pose
de
déterminer
l'existence
éventuelle:
-
d'une
différenciation
génétique
des
populations
des
Pyrénées,
c'est-à-dire
une
évolution,
que
l'on
retrouverait
dans
les
gènes,
diffé-
rente
entre
les
perdrix
des
Pyré-
nées
et
celles
de
plaines.
telle
que
la
Beauce,
par
exemple.
Cette
dif-
férenciation
génétique
pourrait
alors
être
la
conséquence
de
l'iso-
lement
géographique
des
popula-
tions
montagnardes;
-
d'une
pollution
génétique,
c'est-
à-dire
l'introduction
d'allèles
étran-
gers
dans
une
population,
celle-ci
serait
la
conséquence
de
l'introduc-
tion
d'oiseaux
de
repeuplement
issus
d'élevage;
~
d'une
réduction
du
polymor-
phisme
génétique
(voir
définition)
comme
conséquence
de
la
diminu-
tion
des
effectifs.
Description
de
Pétude
Celle-ci
a
porté
sur
5
populations
de
perdrix
jugées
différentes:
Beauce
(BC):
50
individus
prove-
nant
d'un
même
territoire
en
1982.
12
Hautes-Pyrénées
(SS):
25
individus
provenant
de
la
commune
de
Luz-
Saint-Sauveur
en
1983.
Ariège
(}lS).'
9
individus
provenant
de
la
commune
d'Ascou
en
1982.
Pyrénées
orientales
(PO):
Z9
indivi-
dus
provenant
de
la
commune
de
Mantet
en
1982
et
1983.
Elevage
(SB).'
25
individus
prove-
nant
de
l'élevage
de
Saint-Benoist
(78)
de
l'Office
National
de
la
Chasse
en
1982.
Sur
chaque
animal,
des
morceaux
de
muscle
et
de
foie
ont
été
pré-
levés
et
utilisés
pour
cette
étude.
Le
travail
a
porté
sur
l'Z
locus
dif-
férents,
parmi
les
plus
couramment
utilisés
dans
ce
type
d'étude.
Les
techniques
utilisées
pour
sépa-
rer
les
différents
allèles
ne
sont
pas
présentées
ici.
On
pourra
en
trouver
une
description
dans
le
document
intitulé:
0
variation
géographique
de
la
diversité
génétique
chez
la
perdrix
grise
(1985),
disponible
auprès
de
l'auteur
ou
du
Service
Documenta-
tion
de
l'Office
National
de
la
Chasse*.
Plusieurs
méthodes
d'analyse
des
données
ont
été
utilisées:
~
distance
et
identité
génétiques
de
Nei,
0
F.
statistiques
de
Wright,
0
analyse
factorielle
des
corres-
pondances
appliquées
aux
effectifs
alléliques,
aux
effectifs
génotypi-
ques
et
aux
génotypes
individuels.
Leur
description
pourra
également
être
consultée
dans
ce
même
document.
*
G.F.S.
vol.
3,
Mars
1986.
RÉSULTATS
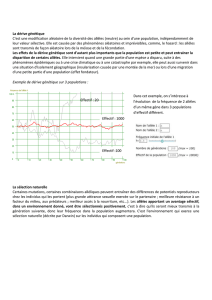
Différence
entre
les
populations
Les
résultats
obtenus
par
les
diffé-
rentes
méthodes
citées
ci-dessus
sont
concordants
entre
eux.
Ils
permettent
de
séparer
le
groupe
de
montagne
pyrénéen
du
groupe
formé
par
la
population
de
Beauce
et
d'élevage.
L'axe
l
qui
posséde
une
inertie
relative
de
77%
sépare
le
groupe
des
Pyrénées,
du
côté
négatif
de
l'axe,
du
groupe
BC-SB
du
côté
positif.
La
position
de
l'échantillon
d'1-\scou
AS,
situé
du
côté
des
Pyrénées,
mais
assez
éloigné
des
deux
autres
échantillons
pyrénéens
PO
et
SS,
est
probablement
due
à
la
taille
de
l'échantillon.
Une
grande
communauté
généti-
que
existe
entre
la
population
d'élevage
de
Saint-Benoist
et
celle
de
Beauce,
suggérant
que
l'essen-
tiel
de
l'élevage
des
perdrix
de
Saint-Benoist
provient
de
la
Beauce.
Toutefois,
la
présence
de
deux
allèles
rares
mais
uniques
a
Saint-
Benoist
pourrait
être
le
témoin
d'un
apport
«étranger»
dans
cet
éle-
vage.
Le
poids
des
différents
gènes
dans
cette
différenciation
inter-
populationnelle
a
été
classé
par
ordre
d'importance.
Une
analyse
factorielle
des
correspondances
montre
que
cette
différenciation
est
essentiellement
due
à
trois
locus
génétiques
(EST-2,
EST
4mu,
1-\AT-2)
sur
les
l'I
étudiés.
Introduction
d'oiseaux
de
repeuplement
L'examen
détaillé
des
résultats
obtenus
au
locus
EST-2
montre
qu'il
existe
dans
l'ensemble
des
cinq
Axe
2
(6%l
4
›
Am
1
(77%)
l"'_\\
ZA
//
\
/
\
Z
//
\
+
///
\
BC
_/
\
SB
+
Fig.
1.
-
Représentation
par
A.T.D.
(
1)
de
la
matrice
des
distances
génétiques.
(1)
A.T.D.:
Analyse
factorielle
d'un
tableau
de
distance:
c'est
la
représentation
dans
un
plan
de
nuage
de
points
formés
à
partir
de
la
matrice
des
distances
entre
uindividusu.

1
l
__
4-
i
1
i
/°'
%_
0
_
,__
i
:›
.Pc
È]
Anne
87
\\\
ra
*r
.
Auele
100
'/
_
Auele
107
Anne
93
Al/èles
ségrégeant
au
locus
EST-2
dans
les
cinq
populations
(surfaces
proportionnelles
aux
fréquences
alléliques).
Fig.
2.
Structure
génétique
au
locus
Est-2
Sur
fond
de
carte:
Répartition
de
la
perdrix
grise
en
France
d'après
Birkan
(1979)
populations,
quatre
allèles
diffé-
rents
a
ce
locus
(fig.
2).
L'allèle
87,
absent
à
Ascou
et
quasi
absent
à
Luz-Saint-Sauveur
(fré-
quence
de
2%),
est
au
contraire
abondant
dans
l'échantillon
de
Beauce
(37%)
et
dans
celui
d'éle-
vage
(35%).
Dans
ce
cas,
la
ques-
tion
est
posée
de
savoir
si
on
peut
le
considérer
comme
marqueur
des
oiseaux
de
plaine?
Un
argu-
ment
de
poids
est
sa
présence
dans
l'échantillon
des
P.O.
(12%)
uniquement
chez
les
individus
qui
pourraient
être
issus
d'un
croise-
ment
avec
un
oiseau
lâché,
d'après
les
informations
communiquées
par
le
technicien
de
l'O.N.C.
ayant
effectué
les
prélèvements
sur
le
terrain.
Si
l'absence
de
cet
allèle
est
confirmée
dans
d'autres
popu-
lations
de
montagne
où
il
n'y
a
eu
aucun
lâcher,
sa
présence
dans
les
P.O.
indiquerait
la
participation
d'oiseaux
de
plaine
ou
d'élevage
à
la
reproduction
du
stock
sauvage,
modifiant
par
la-même
son
origina-
lité.
Ce
point
mériterait
donc
d`être
confirmé
dans
l'avenir.
Niveau
de
polymorphisme
génétique
Une
réduction
de
la
diversité
géné-
tique
est
significative
dans
l'échan-
tillon
d'élevage.
Par
contre,
l'échantillon
de
Beauce
est
parfai-
tement
équilibré,
ce
qui
signifie
qu'il
n'y
a
pas
de
consanguinité.
Dans
les
échantillons
des
Pyré-
nées,
aucune
réduction
n'apparaît
significative
malgré
l'isolement
de
ces
populations
d'altitude
et
l'échantillonnage
qui
comprend,
notamment
dans
les
P.O.
quelques
oiseaux
appartenant
très
probable-
ment
aux
mêmes
compagnies.
Une
analyse
des
génotypes
au
sein
de
ces
compagnies
nous
a
montré
que
des
échanges
d'individus
sont
pos-
sible
entre
elles.
CONCLUSION
Populations
pyrénéennes
Les
populations
pyrénéennes
de
perdrix
se
différencient
des
exem-
plaires
sauvages
de
Beauce
par
la
présence
de
3
allèles
particuliers
et
une
modification
des
fréquences
alléliques
à
4
locus
polymorphes.
Elles
ne
constituent
cependant
pas
un
ensemble
homogène:
chacune
des
populations
échantillonnées
dans
les
Pyrénées
Orientales,
l'Ariège
et
les
Hautes-Pyrénées
manifeste
une
certaine
originalité.
Dans
la
population
des
Pyrénées
Orientales,
une
contamination
géné-
tique
du
stock
naturel
par
des
indi-
vidus
de
repeuplement
a
été
avan-
cée
en
hypothèse.
Population
de
Beauce
Cette
population
apparaît
généti-
quement
équilibrée
puisqu'aucun
locus
ne
montre
de
déséquilibre
significatif.
Population
d'élevage
L'étude
génétique
montre
que
l'essentiel
des
fondateurs
de
cette
population
provenait
principale-
ment
de
Beauce
et
révèle
l'exis-
tence
de
deux
témoins
d'apports
beaucoup
plus
ponctuels
d'oiseaux
étrangers.
La
réduction
significative
de
la
diversité
génétique
observée
uniquement
dans
cette
population
pourrait
s'interpréter
par
une
modi-
fication
des
modalités
d'accouple-
ment
due
aux
conditions
d'élevage.
Cette
hypothèse
serait
à
vérifier
par
une
étude
éthologique.
Apports
de
la
génétique
à
la
gestion
des
populations
de
gibier
L'étude
génétique
réalisée
se
révèle
performante
pour:
-
estimer
le
degré
de
différence
entre
populations
c'est-a-dire
éva-
luer
leur
originalité;
-
estimer
la
diversité
génétique
des
élevages
destinés
au
repeuple-
ment;
-
mettre
en
évidence
une
éven-
tuelle
participation
à
la
reproduc-
tion
en
milieu
naturel
des
individus
de
lâcher,
c'est-à-dire
évaluer
la
contamination
génétique
qu'ils
pro-
voquent
dans
les
populations
sau-
vages;
elle
peut
confirmer,
en
outre,
que
les
échanges
d'individus
entre
compagnies
ne
sont
pas
exceptionnels,
par
la
recherche
des
relations
de
filiation
à
l'aide
des
marqueurs
électrqphorétiques,
à
condition
de
disposer
d'un
mini-
mum
de
polymorphisme.
Les
performances
d'une
telle
ana-
lyse
sont
considérablement
accrues
quand
on
possède
des
informations
biologiques,
écologiques
et
étholo-
giques
sur
les
populations
étudiées.
Nous
avons
le
plaisir
de
remercier
MM.
F.
Biadi,
M.
Catusse,
R.
Gar-
rigues,
R.
Menaut
et
C.
Novoa,
de
l'O.N.C.
ainsi
que
M.
E.
Pelard
et
les
chasseurs
des
communes
de
Luz-Saint-Sauveur,
Ascou
et
Mantet-
Py
pour
leur
contribution
à
la
col-
lecte
des
échantillons.
Cette
étude
a
bénéficié
du
soutien
financier
du
M.U.L.:
convention
218-1982
et
de
l'O.N.C.:
convention
1984
et
1985.
F.
BLANC,
P.
LEDEME,
Ch.
BLANC
Laboratoire
de
Zoogéographie
Université
Montpellier
3
-
BP.
50/13
F
-
34032
Montpellier
Cédex
13
1
/
3
100%