La cellule bacterienne morphologies et structures bacteriennes (word)

UE 8 - De l’agent infectieux à l’hôte
Date : 08/02/16 Plage horaire : 14h-16h
Promo : P2 2015 -2016 Enseignant : JJH
Ronéoiste : CELESTIN Kevin
La cellule bactérienne
Morphologies et Structures bactériennes
I.
Généralités
1.
Qu’est-ce qu’une bactérie
2.
La place des bactéries dans le règne vivant
3.
Organisation des cellules eucaryotes et procaryotes
4.
Morphologies bactériennes
A. Les formes
B. L’arrangement caractéristique
II.
Structures cellulaires
1.
Les composants obligatoires
A. La membrane cytoplasmique
B. Le cytoplasme (cytosquelette, nucléoïde, corps d’inclusions…)
C. La paroi (structure, caractéristiques)
2.
Les structures facultatives
A. La capsule, couche mucoïde, couche S
B. Pili et Fimbriae
C. Les flagelles et la chimiotaxie
D. La spore
E. Les plasmides
a)
Conjugatifs
b)
De résistance
c)
Bactériocines
d)
De virulence
e)
Métaboliques
III.
Taxonomie
1.
Taxonomie moléculaire
2.
Classification médicale
A bon entendeur : il ne va pas insister sur ce point, laissant à ses collègues « l’honneur de le faire »)
NB : Ne pas oublier de ramener une blouse en coton et pas les blouses de service en TP.

I. Généralités
1.
Qu’est-ce qu’une bactérie ?
Observées pour la première fois à la fin du XVIIème siècle (A. Van Leeuwenhoek : « animalcules »), ce n’est
qu’au XIXème siècle qu’on découvre leur rôle dans la fermentation (L. Pasteur) et la transmission de
pathologies (R. Koch, qui a donné son nom aux bacilles de Koch, agents de la tuberculose).
2.
La place des bactéries dans le règne vivant
Procaryotes et eucaryotes font partis du monde du vivant, alors que les virus n’en font pas partis.
Procaryote indique qu’on a à faire à des cellules n’ayant pas de noyaux clairement définis, contrairement
aux eucaryotes chez lesquels on pourra retrouver des êtres unicellulaires ou pluricellulaires.
Parmi les procaryotes, on devra bien distinguer les archées (exemptés de pouvoir pathogène pour l’homme)
des bactéries.
On peut comparer les ordres de grandeur des différents êtres vivants. Certains
parasites (vers intestinaux) peuvent être plus grand que l’homme.
Les bactéries ont une taille de l’ordre du micromètre (de 0,1 à 10 micromètre en
général), équivalente à celle des mitochondries et qui est inférieure à la taille du
noyau d’une cellule eucaryote.
En dessous, on va retrouver les virus qui sont inférieur à 100 nm et en dessous on
retrouvera des protéines (prions…).
Il suffit de regarder
l’arbre du vivant
pour comprendre
que les bactéries
occupent une place
très importante au
niveau de celui-ci.
On retrouvera
énormément de
bactéries pathogènes
ou non pour
l’homme.

3.
Organisation des cellules eucaryotes et procaryotes
En moyenne la taille d’une bactérie va de 0,2 à 2,5 micromètre. On peut tout de même descendre à 0,1
micromètre et monter jusqu’à 100 micromètre voir plus. La taille moyenne d’un eucaryote est comprise
entre 2 et 20 micromètres.
Au niveau de la cellule bactérienne, il n’y a pas de noyau avec une bicouche lipidique qui va déterminer une
structure bien particulière. Néanmoins, le matériel génétique n’est pas libre dans la cellule et va se retrouver
bien localisé.
Classiquement chez les bactéries, on va retrouver un unique chromosome circulaire. Il existe cependant des
bactéries avec plusieurs chromosomes et certaines bactéries peuvent avoir des chromosomes linéaires et non
pas circulaires.
Tous les systèmes
d’empaquetages
endomembranaires n’existent
pas chez les procaryotes
(organites, mitochondries,
lysosomes, golgi, RE).
Néanmoins eucaryotes et
procaryotes possèdent tous une
machinerie cellulaire et
notamment des ribosomes bien
qu’ils se distinguent
structuralement entre procaryotes
et eucaryotes.
4.
Morphologie bactériennes
Les différents aspects concernant la morphologie bactérienne sont essentiels, notamment lorsqu’on veut
faire du diagnostic et de l’identification bactérienne. Il y a plein d’outils possible et utilisables aujourd’hui
(moléculaire…) mais, du moins en clinique, on reste encore avec des critères d’identifications relativement
simples.
A. Les formes
Sphérique (coque) ou bâtonnet (bacille), on distingue principalement 2 formes.
Il existe cependant une
très grande diversité
d’autreformes (spiralées…).

B. L’arrangement caractéristique
Un 2ème niveau d’observation s’intéresse à comment s’organise ces différents types bactériens, ce qui va
permettre de faciliter leur identification. C’est une identification en fonction de la forme et de l’arrangement
caractéristique.
Coques : Cellules individuelles
Paires : diplococcus
Chaines : Streptococcus, Enterococcus, Lactococcus
Amas/grappes : Staphylococcus
Tétrades : Micrococcus
Sarcina : agglomérats de 8 cellules
Ces divers arrangements sont liés au plan de
division cellulaire
Bacilles : Cellules solitaires le plus souvent
Quelque cas de paires ou chaînes : Bacillus megaterium
Rapport longueur/largeur très variable
NB : coccobacilles = très courts + très large aspect de coques
Autres : Vibrions : aspect en bâtonnets incurvés en virgules
Spirilles : bactéries spiralées +/- présence de flagelles (touffes) à 1 ou 2 extrémités
Spirochètes (cas particulier de spirilles) : flagelle interne dans EIM (Leptospira, Borrelia,
Treponema) Actinomycètes : forme un mycélium similaire aux champignon filamenteux.
L’observation au MO (x1000), quoi que peu détaillée, constitue la 1ère étape de l’identification (schéma ci-
dessus à bien connaître).
IV.
Structures cellulaires
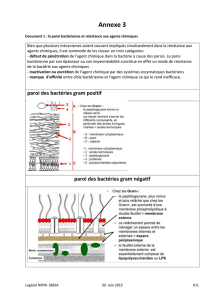
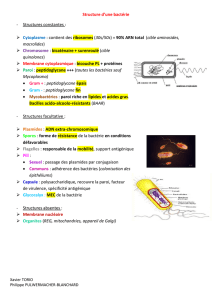
Toutes les bactéries vont au moins disposer d’une paroi bactérienne (composée de peptidoglycanes),
puis d’une membrane cytoplasmique entourant donc un cytoplasme contenant un nucléoïde.
Effectivement, en l’absence de noyau c’est dans cet espace que s’organise le matériel génétique (ADN,
ARN, protéines). Des ribosomes et des corps d’inclusion (faisant office de réserve énergétique) sont aussi
retrouvés. L’ensemble de ces éléments représentant les éléments communs obligatoires.

D’autres structures comme la capsule, la couche S, ou des systèmes entraînant la formation d’une sorte de
mucus, mais aussi les flagelles, vont faire parties des structures facultatives des bactéries.
Membrane cytoplasmique : barrière perméable sélective, limite mécanique de la cellule, transport des
éléments nutritifs et des déchets, localisation de plusieurs processus métaboliques (respiration,
photosynthèse), détection de signaux de l’environnement pour le chimiotactisme.
Espace péri-plasmique : Contient des enzymes hydrolytiques et les protéines de liaison nécessaires à la
capture et la transformation nourriture.
1.
Les composants obligatoires
A. La membrane cytoplasmique
Elle ressemble parfaitement à la membrane cytoplasmique d’une cellule eucaryote et est constituée :
-
D’une bicouche lipidique (têtes polaires, queues apolaires)
-
De protéines intrinsèques et extrinsèques
-
Sur la face externe, des sucres sont rattachés soit aux protéines soit aux lipides membranaires.
Ainsi, comme chez les eucaryotes, cette membrane est asymétrique et polarisée.
Dans beaucoup d’ouvrages on indique que les bactéries, pour augmenter leur espace d’échange avec le
milieu environnant sont capables de former des replis membranaires (sortes d’invaginations membranaires)
qui semblent indiqués qu’à certains endroits on peut avoir des extensions associées à des fonctions bien
particulières. Les dernières études semblent montrer que ce sont des artéfacts expérimentaux. En réalité,
c’est lorsqu’on fixe les bactéries et qu’on essaye de les observer qu’on observe ces structures mais que
naturellement elles n’existeraient pas. Toutes ces structures qu’on appelle des mésosomes, des lamelles ou
des tubes qui seraient liées à des invaginations membranaires plus ou moins complexes seraient des
artefacts qui n’existeraient pas en réalité.
La bicouche lipidique doit avoir une certaine fluidité. Chez les eucaryotes, cette fluidité peut être modulée
en fonction de la température de l’environnement par des stérols (cholestérol). Les bactéries n’ont quasiment
pas de cholestérol à quelques exceptions près. Cependant les hopanoïdes, un type particulier de stérol, vont
venir jouer le même rôle de modification de la fluidité membranaire chez les bactéries.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%