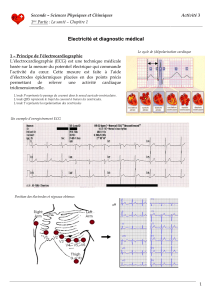
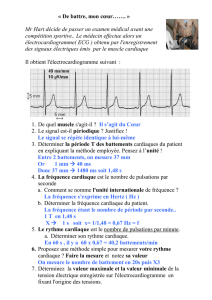
Moinnereau_Marc_Antoine_MScA_2017.pdf (7.970Mb)



UNIVERSITÉ DE SHERBROOKE
Faculté de génie
Département de génie électrique et de génie informatique
ENCODAGE D’UN SIGNAL AUDIO DANS
UN ÉLECTROENCÉPHALOGRAMME
Mémoire de maîtrise
Spécialité : génie électrique
Marc-Antoine MOINNEREAU
Jury : Éric PLOURDE (directeur)
Kevin WHITTINGSTALL (co-directeur)
Denis GINGRAS
Yves BÉRUBÉ-LAUZIÈRE
Sherbrooke (Québec) Canada Mai 2017


RÉSUMÉ
Les interfaces cerveau-machine visent à établir un lien de communication entre le cer-
veau et un système externe à ce dernier. Les électroencéphalogrammes (EEG), dans ce
contexte, ont l’avantage d’être non invasifs. Par contre, l’information sensorielle qui se
retrouve dans un signal EEG est beaucoup moins ciblée que dans un signal neuronal
acquis par une méthode invasive. De plus, étant donné que le cortex auditif est situé
dans des repliements du tissu cortical, les neurones qui déchargent, suite à un stimulus
auditif, sont parallèles à la surface corticale sur laquelle les EEG sont enregistrés. Par
conséquent, l’information auditive qui se retrouve dans le canal EEG situé vis-à-vis du
cortex auditif est faible. L’objectif principal de ce projet de recherche consiste donc à
étudier la répartition de l’information auditive dans l’ensemble des canaux EEG. Pour ce
faire, nous utilisons deux approches. Dans la première, nous tenterons d’estimer l’activité
corticale sous-jacente à partir des signaux EEG en utilisant un modèle de couplage bande
fréquence. En effet, certaines bandes de fréquences sont des bons prédicteurs des décharges
neuronales. Cependant, cette approche n’a pas été validée pour le système auditif, nous
confronterons donc l’estimation obtenue à une autre estimation en ayant recours à un
modèle spécialisé pour l’encodage du signal de parole faisant appel aux processus ponc-
tuels. Ce modèle prend en compte les dynamiques intrasèques des neurones et également
des propriétés spectrotemporelles du stimulus d’entrée. Dans la seconde approche, nous
étudierons la possibilité de classifier 3 voyelles (a, i et u) en fonction du nombre de canaux
EEG utilisés ainsi que leur répartition sur le cuir chevelu. Nous aurons recours, pour cela,
à un réservoir de neurone à décharge récurrent activé en entrée par les données EEG. Les
résultats démontrent que l’information auditive se retrouve en fait dans l’ensemble des
canaux EEG et qu’elle n’est pas confinée à un nombre restreint d’électrodes. Il est égale-
ment montré que lorsque l’on utilise les 64 électrodes que comporte l’EEG pour classifier
les 3 voyelles, on obtient une classification de l’ordre de 80%, mais aussi qu’un nombre
limité de 10 électrodes suffit pour obtenir une classification satisfaisante et, qu’en plus, la
position de ces électrodes sur le cuir chevelu est peu importante.
Mots-clés : Électroencéphalogramme (EEG), Encodage neuronal, Modèle linéaire gé-
néralisé, Données spatio- et spectro-temporelles, Réservoir, Réseaux de neurones
récurrents, Apprentissage machine, Cortex auditif
i
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
1
/
84
100%