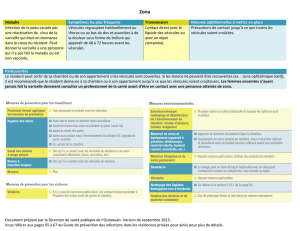
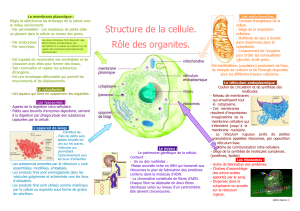
Système endomenbranaire

Système endomembranaire
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
2 compartiments
-Intra cellulaire
-Extra cellulaire
Systèmes endomembranaires :
Compartiments membranaires intracellulaires fonctionnant en relation
avec la membrane plasmique.
Réticulum endoplasmique
granuleux
Réticulum endoplasmique
lisse
Appareil
de Golgi
Vésicules
Lysosomes
Systèmes endomenbranaires : Compartiments membranaires intracellulaires fonctionnant
en relation avec la membrane plasmique.
Définitions et généralité
!
Un organisme vivant est constitué par des cellules, et que ces cellules ont fabriqués de la
MEC qui s'est ensuite accumulé entre les cellules.
On a 2 compartiments, intra et extra-cellulaire et dans la cellule on a aussi 2 territoires,
celui qui va rester dans la cellule, et un qui va pouvoir produire des protéines qui vont
pouvoir sortir de la cellule et fabriquer la MEC par exemple.
C'est le système endomembranaire, qui est constitué par des compartiments
membranaires en continuité dans le temps et dans l'espace est en continuité avec la
membrane plasmique.
Toutes ces cavités, qui ont une lumière blanche, sont en continuité avec le milieu extra-
cellulaire. Et les membranes sont en continuité avec la membrane plasmique dans le
temps et dans l'espace.
On a des réticulums, des lisses et granuleux, l'appareil de Golgi, et différents types de
vésicules (donc les lysosomes).
On a des compartiments avec des tailles, des formes et des positions variables et
différentes. Ils sont tous entourés par une membrane qui est une bicouche
phospholipidique. En continuité dans le temps et dans l'espace avec la membrane
plasmique.
Fonctions du système endomembranaire.
Les fonctions de ce système sont très importante dans la synthèse de ce milieu extérieur,
car le milieu extra-cellulaire est en continuité avec les lumières dans lesquelles sont
produites dans protéines, des glycoprotéines en particulier.
Système endomembranaire : Fonctionnements :
- En perpétuel dynamique (synthèses, échanges entre les compartiments, dégradation)

- Echanges avec la membrane plasmique et le milieu extérieur (vésicules, exocytose,
endocytose)
Système endomembranaire : Fonctions :
- Synthèses
- Métabolismes
- Catabolismes
- Transports intracellulaires, sécrétions
Il va aussi synthétiser les membranes, la membrane plasmique, et celles des différents
membranes internes de la cellule.
Phénoméne d'exocytose
Phénomènes de phagocytoses, englobement de grosses particules par une cellule qui se
fait pas un système d'englobement tapissé par la membrane plasmique et celle des
phagosomes. Cette phagocytose est impliqué dans la destruction, et en particulier dans le
renouvellement de la MEC.
Il y a des phénomènes de métabolisme: par le biais d'endocytose des particules contenant
du cholestérol. Qui est ensuite impliqué dans la fabrication des membranes.
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
Appareil de
Golgi
Tri et adressage
Modifications moléculaires
post-traductionnelle et autres
Formation de vésicules
Réticulum endoplasmique
lisse
Réticulum endoplasmique
granuleux
Synthèse de lipides
et de stéroïdes
Stockage et
libération du calcium
Autres fonctions
Synthèse
protéïque
(traduction)
Lysosome
Hydrolyse
Dégradation
Grain de
Sécrétion
Vésicule de
transport
Sécrétion
régulée
Sécrétion
constitutive
Systèmes endomembranaires
Lysosomes
Peroxysomes
Protéasome
Il y a des transports, des mouvements, et grâce à ces mouvements des orientations
certaines vésicules vont allé a telles ou telles pôles de la cellules, on va avoir une
compartimentation cellulaire.
On a les vésicules de transport qui assure la sécrétion, les grains de sécrétions qui
assurent la sécrétion régulée, la mission est de libérer des composés par exocytose.
phénomènes d'exocytoses régulables
Les lysosomes et peroxydoses comportent des cytomembranes, qui sont des enzymes de
dégradation, et le protéases mais nʼa pas de cytomembrane.

Cavités : citernes intercommunicantes délimitées par des endomembranes
composéesdʼune bicouche de phospholipides, elles sont intercommuniquantes.
(Reticulum, appareil de Golgi … )
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
CAVITES :
Citernes intercommunicantes délimitées par des endomembranes
5pWLFXOXPV$SSDUHLOGH*ROJL««
Bicouche de phospholipides
Protéines et sucres associés
Systèmes endomembranaires ± structure
VESICULES :
Systèmes clos. Une endomembrane entourant un contenu
/\VRVRPHV(QGRVRPHV*UDLQVGHVpFUpWLRQ««
Bicouche de phospholipides
Protéines et sucres associés
Vésicules : système clos, rond, sphérique, une cytomembrane entourant un contenu.
Selon le compartiment, les protéines seront de nature différentes (lysosome, endosome,
grains de sécrétion … )
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
Synthèses, métabolismes et
transports intracellulaires dans le cytoplasme
(Pancréas exocrine)
Réticulum endoplasmique
granuleux (synthèse protéique)
Grains de sécrétion
(grains de zymogène)
(Stockage et exocytose)
Appareil de Golgi
(métabolisme et formation
de vésicules)
Exocytose
Ces composants sont organisés spatialement dans des dispositifs spécifiques de chaque
population cellulaire, que ce soit par la nature des contenus ou dans le positionnement a
lʼintérieur de la cellule. La cellule pancréatique exocrine a pour fonction de produire des
enzymes libérés dans le tube digestif, ce sont des protéines stockées dans des grains de
sécrétion (grains de zimogène) qui stockent des enzymes et les libèrent par exocytose
dans le milieu extérieur. Le REG va produire ces enzyme, lʼAG va empaqueter ces
enzymes.
La cellule du pancréas exocrine (exocrine : libération dans une cavité en contact avec le
milieu exterieur) : elle a pour but de fabriquer des enzymes de la digestion qui vont être

libérées dans le tube digestif, et qui vont servir a la digestion. Les enzymes sont des
protéines, la cellule est donc spécialisée dans la fabrication de protéines, a destination du
milieu extra-cellulaire. On a donc toute la machinerie nécessaire a la production d'une
grande quantité de protéines: REG ou se fait la traduction, un AG territoire en interface
entre les réticulum et ses dérivés, orientation des composés présents dans la cellule, des
modifications des protéines et la formation de vésicules. Les enzymes sont emballées
sous forme inactive dans des vésicules, dans des grains de sécrétion (grains de
zymogène dans le pancréas) .Quand on mange signal qui explique a la cellule
pancréatique qu'il faut qu'il libére les enzyme → exocytose.
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
Réticulum
endoplasmique
Granuleux (REG)
Réticulum
endoplasmique lisse (REL)
I- Le réticulum endoplasmique
!
Présents dans (tous) les types cellulaires
50% des membranes de la cellule
10 à 30 fois la surface de la membrane plasmique
Abondansce du REL et du REG très variable selon les besoins et types cellulaire et en
fonction de lʼactivité de la cellule
Citernes intercommunicantes délimitées par une bicouche phospholipidique
- plus mince que la membrane plasmique (pas de cell coat)
- memes catégories de lipides
- protéines (dont enzymes ++)
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
Réticulums endoplasmiques
Présents dans (tous) les types cellulaires
50% des membranes de la cellule
10 à 30 fois la surface de la membrane plasmique
Abondance du REL et du REG très variable selon besoins et
W\SHVFHOOXODLUHVHWHQIRQFWLRQGHO¶DFWLYLWpGHODFHOOXOH
Citernes intercommunicantes délimitées par une bicouche
phospholipidique
±plus mince que la membrane plasmique (pas de cell coat)
±Mêmes catégories de lipides
±Protéines (dont enzymes ++)
1- Caractéristiques générales
!
Le réticulum, on a deux types, le lisse et le granuleux. Le réticulum est une citerne bordée
par la bicouche phospholipidique plus mince que celle de la membrane plasmique mais
avec les mêmes constituants, les mêmes catégories de lipides mais en proportions
différentes, des protéines y sont enchâssées, dont certaines seront des transporteurs et
d'autres des enzymes.

Ces citernes sont plus ou moins abondantes selon les besoins de la cellule. On a des
cellules riches en REL ou en REG; on a des régulations de quantité dans le temps.
Lymphocyte = cellule au repos, qui n'as pas d'activité importante au repos. Si ces
récepteurs reconnaissent un antigène, il s'active, on a un transformation, et une
augmentation du REG pour fabriquer des protéines pour se défendre.
Les cellules seront soit actives, soit sous forme de cellule mémoire au repos (avec un RE
de faible quantité )
En moyenne ces réticulums représentent une part importante des membranes cellulaires
(50% environ), c'est 10 à 30 fois la surface de la membrane plasmique.
Ces réticulums se répartissent dans certains territoires, la localisation est différente selon
les cellules.
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
Les réticulums dans
la cellule pancréatique exocrine et le neurone
Réticulum endoplasmique
granuleux
Grains
de zymogène
Appareil
de Golgi
Axone (réticulum lisse)
Corps cellulaire
(réticulum granuleux et
lisse)
Pancréas exocrine Neurone
Exemple du pancréas exocrine: cellule avec bcp de REG, qui se retrouve en position infra
et péri nucléaire.
2011- 2012
UE2 - La cellule et les tissus
Systèmes endomembranaires et trafics intracellulaires. Pr B.Bloch
Les réticulums - microscopie optique
Réticulum endoplasmique granuleux
(Corps de Nissl; neurone)
Réticulum endoplasmique lisse
(Corticosurrénale; cellules endocrines
SURGXFWULFHVG¶KRUPRQHVVWpURwGHV
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
1
/
34
100%