l`identite biologique des organismes et - sommaire SVT-fxm

1/7 1S SVT– cours vision 1
Cours vision 1 : structure de l’œil, cristallin, rétine, etc.
La vision est un des sens les plus importants parmi les 5.
Nous allons voir comment ça marche.
I. La structure de l’œil.
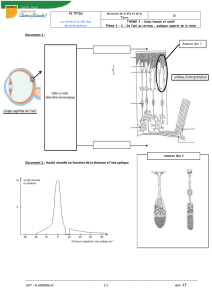
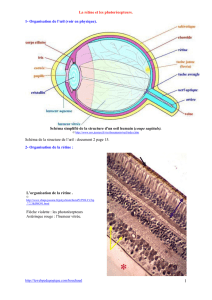
Lors de la dissection, on constate que l’œil est constitué d’une enveloppe délimitant
une cavité, à l’intérieur de laquelle se trouve un liquide gélatineux : l’humeur vitrée.
L’enveloppe de l’œil est composée de plusieurs couches :
La sclérotique (blanche),
La choroïde (noire)
La rétine (rose).
Au niveau de la pupille, la sclérotique est remplacée par la cornée, la choroïde par
l’iris.
Le cristallin se présente sous forme d’une petite structure sphérique dure et
transparente.
Les structures traversées par la lumière sont :
La cornée,
L’humeur aqueuse,
Le cristallin,
L’humeur vitrée.
Ces quatre structures sont incolores et parfaitement transparentes.
La sclérotique, couche épaisse et blanche (rose vif ici à cause de la coloration),
constitue l’enveloppe externe de l’œil. Non vascularisée, elle donne une certaine
résistance à cet organe.
À l’avant, la sclérotique devient transparente et constitue la cornée.
La choroïde, pigmentée (noire en coupe), est richement vascularisée, elle assure la
nutrition des cellules de l’œil.
À l’avant elle se prolonge par l’iris.
La rétine, couche interne de l’œil, rose pâle, s’interrompt au niveau de la pupille.
II. Le cristallin une lentille perfectionnée et vivante.
A. Organisation.
Doc. 1 : La fonction du cristallin est de laisser passer la lumière venant de l’extérieur
de l’œil, en assurant une convergence des rayons lumineux. Le degré de vergence du
cristallin est variable, ce qui permet l’accommodation, c’est-à-dire la mise au point
d’une image nette sur la rétine lors de la vision d’objets rapprochés.
Doc. 2 :
– Forme parallélépipédique, en « ruban », perpendiculaire au trajet des rayons
lumineux
→ pas de dispersion ;

2/7 1S SVT– cours vision 1
– Cellules sans organites, sans noyau, protéines en réseau cristallin, cytoplasme sous
forme de « gel » → transparence ;
– Nutrition à partir de nutriments solubles transmis par diffusion (pas de capillaires
sanguins) → transparence.
B. Le cycle des cellules cristallines.
Doc. 2 et 3 : Les cellules du cristallin ont une vie qui repose sur les mêmes principes
que ceux qui régissent la vie de toute cellule : métabolisme exploitant des nutriments
fournis aux cellules, mitoses, expression d’un programme génétique, renouvellement
du contenu cellulaire. Cependant, les modalités de ce fonctionnement sont très
particulières :
– l’apport des nutriments ne se fait pas par les capillaires sanguins mais par simple
diffusion à partir de l’humeur aqueuse ;
– l’information génétique est exploitée au début de la vie cellulaire puis le noyau est
éliminé, ce qui explique la transparence du contenu cellulaire mais aussi les capacités
limitées qu’ont ces cellules pour se réparer ;
– enfin, les cellules du cristallin se distinguent par leur exceptionnelle longévité.
C. Les anomalies du cristallin.
Doc. 4 : La presbytie s’explique par un vieillissement normal des cellules du
cristallin.
En effet, l’accommodation nécessite une très bonne élasticité des cellules du
cristallin.
Comme ces cellules ne sont pas renouvelées, il est compréhensible que cette élasticité
diminue avec l’âge.
Les cellules du cristallin ne se renouvellent pas et perdent rapidement leur noyau :
elles ne pourront donc plus exploiter l’information génétique pour remplacer des
protéines déficientes par exemple. Ainsi, contrairement aux autres tissus, les lésions
subies par le cristallin et qui peuvent altérer sa transparence sont difficilement
réparées.
D. Bilan.
Les cellules du cristallin sont dépourvues de noyau et d’autres organites et leur
cytoplasme est parfaitement transparent. Elles laissent donc passer la lumière. Elles
ont une forme très allongée et sont agencées en un ensemble biconvexe qui assure la
convergence des rayons lumineux. Leur élasticité permet une modification de la
forme générale du cristallin et donc l’accommodation.

3/7 1S SVT– cours vision 1
III. La rétine.
A. Organisation.
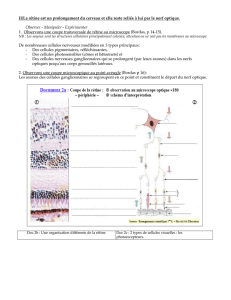
Doc. 1 et 2 : Les cellules photoréceptrices de la rétine sont les cônes et les bâtonnets.
Elles sont localisées dans la couche cellulaire la plus externe de la rétine.
Doc. 1 : Chaque photorécepteur est lié à un neurone bipolaire, lui-même connecté à
un neurone ganglionnaire dont le prolongement cytoplasmique constitue une fibre du
nerf optique.
B. Les photorécepteurs : des cellules nerveuses.
Doc. 3 Cette étude montre que le cône réagit à un stimulus lumineux par une
variation du potentiel électrique de sa membrane. Cette cellule est donc sensible à la
lumière.
De plus, on constate que plus l’intensité du stimulus est importante, plus la variation
du potentiel électrique l’est également, jusqu’à une valeur qui semble être maximale.
Doc. 1 à 3 : La rétine est un tissu nerveux car elle est constituée d’un ensemble de
cellules nerveuses interconnectées. Elle possède la propriété de réagir par une
variation de potentiel électrique à une stimulation extérieure (ici la lumière), ce qui
est une propriété caractéristique d’un récepteur sensoriel.
Les cellules photoréceptrices de la rétine constituent l’assise cellulaire périphérique
de cette fine membrane. Ce sont de petites cellules, allongées et très fines, renfermant
un pigment photosensible et réagissant à un stimulus lumineux par une variation de
potentiel électrique. Ces cellules sont connectées à des neurones qui forment les
fibres du nerf optique.
C. Les deux types de récepteurs.
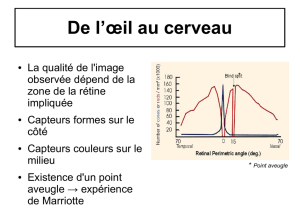
1. Vision centrale, périphérique et sensibilité des récepteurs.
Doc. 1 et 2 : La vision centrale se fait au niveau de la fovéa, essentiellement garnie
de cônes. Ceux-ci étant peu sensibles, elle nécessite un plus fort éclairement (vision
diurne). Du fait de la densité maximale des cônes, l’acuité visuelle est excellente dans
ces conditions. Comme cette vision implique les différentes catégories de cônes, la
vision des couleurs est possible.
La vision périphérique fait, quant à elle, appel aux portions de rétine situées en
dehors de la fovéa, plus riches en bâtonnets qu’en cônes. Du fait des caractéristiques
des bâtonnets, la vision périphérique est donc possible même si l’éclairement est
faible (vision nocturne) ; en revanche, la vision périphérique est peu précise. Les
bâtonnets ne permettent pas de distinguer les couleurs (une seule sorte de
photorécepteurs) et la vision s’effectue donc en « nuances de gris ».
Doc. 1 et 2 : La lecture nécessite un bon éclairement puisqu’elle est réalisée par les
cônes (photorécepteurs ultra-majoritaires dans la fovéa), qui sont peu sensibles
(moins que les bâtonnets). Or, cette zone où la densité des cônes est maximale est de

4/7 1S SVT– cours vision 1
petite taille : pour que l’image du texte se forme continuellement sur cette zone, l’oeil
doit suivre la ligne, mot à mot en permanence, de façon très précise.
2. La vision des couleurs.
Doc. 3 : En effet, chez une personne qui ne peut pas produire d’opsine sensible au
vert, une radiation lumineuse verte ne pourra être perçue que par les cônes sensibles
au rouge. Comme le montre les spectres d’absorption, une radiation verte (ex. 525
nm) sera absorbée avec la même intensité par les cônes sensibles au rouge qu’une
radiation rouge (ex. 625 nm). Ces deux couleurs seront donc perçues de la même
façon.
Doc. 3 : Il existe d’autres anomalies de la vision des couleurs. Par exemple, d’autres
formes de daltonisme sont dues à l’absence d’opsine sensible au rouge ou à l’absence
d’opsine sensible au bleu (beaucoup plus rare). Dans tous ces cas, l’ensemble des
nuances colorées ne sont restituées qu’à partir de deux sources au lieu de trois : de ce
fait, certaines couleurs sont perçues de façon identique et sont donc confondues.
L’achromatopsie, due à l’absence totale d’opsines, existe mais est rarissime : dans ce
cas, le sujet ne voit le monde qu’en nuances de gris.
D. Densité et acuité.
Analysez ce document et expliquez pourquoi l’acuité visuelle est maximale au
« centre ».
Les cônes et les bâtonnets ne se répartissent pas de façon identique :
- la densité de cônes est maximale au niveau de la fovéa et diminue très rapidement
de part et d’autre de celle-ci ;
- inversement, les bâtonnets présentent une densité plus importante dans la rétine
périphérique que dans la fovéa.
Comme cela a été vu dans l’activité précédente, cônes et bâtonnets sont absents du
point aveugle, point de départ du nerf optique.
La vision des couleurs est maximale dans la zone centrale qui se projette dans la zone
centrale de la rétine. On peut donc associer vision des couleurs et cellules
photoréceptrices présentes dans cette zone : les cônes sont responsables de la vision
des couleurs.
La vision des couleurs est absente en périphérie et les cellules photoréceptrices de la
rétine périphérique sont des bâtonnets : ils sont donc responsables de vision en teintes
de gris.
L’acuité visuelle est maximale dans la zone de la fovéa : le niveau de précision avec
lequel sont perçus les objets est donc maximal dans la zone centrale du champ de
vision.

5/7 1S SVT– cours vision 1
E. Bilan.
Les cônes sont des photorécepteurs relativement peu sensibles à la lumière mais
permettant une vision très précise. L’existence de trois types de cônes, renfermant des
pigments différents, permet une vision trichromatique des couleurs. La très grande
densité des cônes dans la fovéa (région centrale de la rétine), permet la vision centrale
diurne (exigeant un bon éclairement) que l’on utilise par exemple au cours de la
lecture.
Les bâtonnets sont beaucoup plus sensibles à la lumière, mais ils ne permettent de
percevoir ni les détails ni les couleurs. La prédominance des bâtonnets dans la région
périphérique de la rétine permet néanmoins une vision dans de faibles conditions
d’éclairement.
IV. La vision des couleurs et liens de parenté.
A. La famille multigénique des opsines.
Doc. 1 et 2 : Les pigments rétiniens (opsines) présentent de grandes similitudes. La
séquence de l’opsine sensible au bleu présente plus de 40 % d’identité avec la
séquence de l’opsine sensible au rouge. Cette dernière possède plus de 95 %
d’identité avec celle de l’opsine au vert. Ces protéines sont bien entendues codées par
des gènes. Il serait très surprenant que de telles similitudes soient dues au simple
hasard. Il est beaucoup plus logique de faire l’hypothèse que ces gènes ont en fait une
origine commune, ce qui expliquerait alors l’existence de telles similitudes.
Les différences entre ces gènes peuvent alors facilement s’expliquer : l’ADN étant
relativement instable (voir Partie 1 chapitre 2), des mutations ponctuelles
s’accumulent au cours du temps, ce qui différencie peu à peu ces gènes, initialement
identiques.
 6
7
6
7
1
/
7
100%