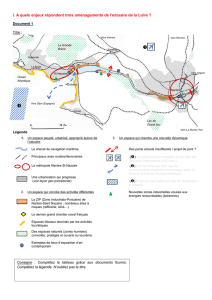
Quel référentiel de fonctionnement pour les

UNIVERSITE BORDEAUX I
ECOLE DOCTORALE SCIENCES DU VIVANT-GEOSCIENCES-SCIENCES DE L’ENVIRONNMENT
N° d’ordre : 2890
E. Rochard
C. Genty
C. Genty
Cemagref
Cemagref
E. Rochard
C. Genty
C. Genty
Cemagref
Cemagref
T H E S E
pour obtenir le grade de
DOCTEUR DE L'UNIVERSITE BORDEAUX I
Discipline : OCEANOGRAPHIE
« Quel référentiel de fonctionnement
pour les écosystèmes estuariens ? »
Le cas des cortèges de poissons fréquentant
l’estuaire de la Gironde
présentée et soutenue publiquement
par
Jérémy LOBRY
Le 19 novembre 2004
_______
Devant le jury composé de :
M. Pierre CHARDY (Professeur, Bordeaux I), président
Mme Maria-José COSTA (Professeur, Université de Lisbonne), rapporteur
M. Bernard HUGUENY (Chargé de Recherches, IRD), rapporteur
M. Didier GASCUEL (Professeur, ENSAR), examinateur
M. Eric ROCHARD (Directeur de Recherches, Cemagref), directeur de thèse
M. Pierre ELIE (Directeur de Recherches, Cemagref), directeur de thèse


Ce sont rarement les réponses qui apportent la vérité, mais l'enchaînement des questions
(Daniel Pennac – La fée carabine)
i

« QUEL REFERENTIEL DE FONCTIONNEMENT POUR LES ECOSYSTEMES ESTUARIENS ? »
LE CAS DES CORTEGES DE POISSONS FREQUENTANT L’ESTUAIRE DE LA GIRONDE
ii

REMERCIEMENTS
Belle habitude, quoique assez étrange finalement, que de commencer une thèse par les
remerciements. J’ai longtemps attendu ce moment. D’abord parce qu’il est synonyme de la
l’aboutissement du travail. Ensuite parce qu’il me permet enfin de remercier toutes les personnes
qui, par leur présence, leurs conseils, leur patience, leur écoute, leur amitié ou leur amour, m’ont
permis de tenir jusque là et de présenter, enfin, mon travail devant un jury.
Commençons par lui, justement. Jusqu’ici, je n’avais pas eu la chance de rencontrer M. Bernard
Hugueny et Mme Maria-José Costa. Qu’ils soient remerciés pour l’honneur qu’ils m’ont fait en
jugeant mon travail et en « rapportant » mon mémoire. Je remercie tout particulièrement Mme
Costa qui a accepté de le faire dans une langue qui n’est pas la sienne. Pour cela :
MUITO
OBRIGADO
!
Merci aussi à M. Pierre Chardy que j’ai déjà eu la chance de croiser à quelques reprises lorsqu’il a
répondu avec gentillesse à mes invitations. Ses conseils, ses analyses et ses critiques furent à
chaque fois constructifs et précieux. Enfin, dernier membre de cette commission, et non des
moindres, je souhaite exprimer toute ma reconnaissance à M. Didier Gascuel. C’est à cause de lui
que tout a commencé. Didier a été le premier à me donner ma chance. Il fut mon professeur puis
mon responsable de stage. Son dynamisme et son sens didactique m’ont donné envie de suivre
cette voie. J’espère pouvoir encore travailler avec lui le plus souvent possible et je le remercie
chaleureusement d’avoir accepté de faire partie de ce jury.
En paraphrasant Gisèle Parfait (Spot, Mai 2004), je dirais que faire une thèse est une véritable
« aventure humaine, un parcours du combattant semé de rencontres ». Parmi ces rencontres,
celles de Pierre Elie et Eric Rochard furent évidemment déterminantes. Je souhaite leur
exprimer ma plus profonde gratitude pour m’avoir accordé leur confiance jusqu’au bout. J’espère
en avoir été digne. Merci à toi Pierre pour ton appui indéfectible même lorsque nous n’étions pas
forcément d’accord et pour m’avoir fait partager ton expérience des milieux estuariens et de la
recherche en général. Merci de m’avoir donné la possibilité de venir faire cette thèse au
Cemagref. Thèse qui n’aurait probablement pas été la même sans le soutien constant d’Eric
Rochard. Ses qualités scientifiques et humaines, sa disponibilité, son ouverture d’esprit, son sens
de l’encouragement et cette faculté irremplaçable à me suggérer tout un tas d’idées en me
laissant croire qu’elles venaient de moi m’ont conduit jusqu’ici. Le grand plaisir que j’ai eu à
travailler avec lui allié à une volonté constante d’essayer de ne pas le décevoir ont été un moteur
précieux tout au long de ma thèse.
J’ai eu la chance de travailler durant ces quelques années de thèse au sein de l’unité Ressources
Aquatiques Continentales du Cemagref. Merci à Paul Gonthier de m’y avoir accueilli et de m’avoir
initié aux conventions avec le ministère, aux réponses aux appels d’offre et à la tenue de délais
serrés (« au fait, Jérémy, on a un rapport à rendre pour hier… ! »).
4 ans, 4 ans attente d’une date propice à la soutenance comprise : une thèse, c’est long ! Ca paraît
tout de suite moins long lorsque l’on est bien entouré. Merci à Mario Lepage qui s’rait un peu du
genre à avoir la bonne humeur communicative pour m’avoir toujours associé au fonctionnement de
l’équipe, pour m’avoir toujours soutenu et avoir généreusement partagé ses connaissances sur
l’écologie des espèces et son expérience de terrain, pour ses anecdotes, pour m’avoir initié au
Pitchak et au badminton ; à Patrick Lambert, mon bizuth (ce fut un honneur), pour ses bons mots,
ses « judicieux » conseils, ses « pertinentes » questions y compris trois jours avant la soutenance
et aussi pour toutes ces discussions scientifico-philosophiques (désolé de t’avoir empêché de
iii
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
1
/
230
100%