biologie du fdg et des autres traceurs émetteurs de positons utilisés

J.Ph. VUILLEZ
4Revue de l'ACOMEN, 1998, vol.4, n°1
Lutilisation diagnostique des radiopharmaceutiques en
cancérologie a jusquici surtout concerné la détection des
tumeursmais il savère de plus en plus quelle portera dans
lavenir sur la caractérisation tumorale in vivo. La détec-
tion (= affirmer la présence dun cancer) se situe à diffé-
rents niveaux : diagnostic positif, bilan dextension, mala-
die résiduelle, détection de récidives. Elle repose sur la
mise en évidence, au moyen dune molécule marquée ap-
propriée, de caractéristiques biochimiques ou fonction-
nelles spécifiques, soit des cellules tumorales elles-mê-
mes, soit de la réaction de lorganisme à la présence dune
tumeur (cas de la scintigraphie osseuse). Les caractéristi-
ques ciblées sont lexpression de récepteurs, des antigè-
nes dits "tumoraux", certains processus biochimiques (ddp
trans-membranaire), certaines propriétés métaboliques.
Les radiopharmaceutiques qui permettent ce ciblage spé-
cifique des tumeurs permettent donc par nature une ca-
ractérisation tumorale plus ou moins fine. Lévolution
actuelle de la cancérologie repose en grande partie sur les
possibilités et les techniques de cette caractérisation tu-
morale, aboutissement du progrès des connaissances en
Biologie Cellulaire et de la compréhension des processus
de carcinogenèse en sappuyant sur des notions telles
que le pouvoir de prolifération, la chimiosensibilité, la ra-
diosensibilité, lagressivité et pouvoir métastasiant, le de-
gré de différenciation, le statut hormonal et plus largement
lexpression de récepteurs (hormonaux, mais aussi pour
les cytokines, les facteurs de croissance), lexpression
dantigènes, la viabilité cellulaire, l état métabolique, lex-
pression doncogènes et antioncogènes Létude de ces
paramètres a (au moins) une triple finalité : lévaluation du
pronostic, l appréciation de lefficacité du traitement et le
choix des modalités de traitement, que la caractérisation
de la tumeur permet denvisager de façon rationnelle et
adaptée. Elle passe le plus souvent par des méthodes
danalyse tissulaire, qui nécessitent de disposer dun pré-
lèvement, biopsique ou chirurgical : immunohistochimie,
études de récepteurs, études de biologie moléculaire, do-
sages denzymes
Les méthodes dinvestigation in vivo non invasives per-
mettent de surveiller ces différents paramètres sans inter-
vention sanglante, et ont en outre lavantage de permettre
lanalyse simultanée de toutes les localisations tumorales
chez un patient, alors que les biopsies sont nécessaire-
ment limitées à un ou quelques sites seulement. Cest une
voie de développement potentiellement très féconde de la
Médecine Nucléaire. Les études métaboliques en particu-
lier connaissent un grand succès, du fait quelles permet-
tent de très bien appréhender le degré de malignité à tra-
vers des aspects fondamentaux de la prolifération cellu-
laire comme la consommation énergétique accrue, laug-
mentation de la synthèse protéique ou encore la synthèse
dacides nucléiques. Les molécules concernées par ces
processus sont en général de petites molécules (hexoses,
acides aminés, bases puriques et pyrimidiques) et leur
marquage par des émetteurs gamma, sans être nécessaire-
ment utopique (1-3), est difficile sans dénaturer les pro-
priétés de ces molécules. Ceci explique lattrait du mar-
quage par des isotopes émetteurs de positons, qui néces-
sitent peu ou pas de modifications chimiques.
A. Le fluoro-déoxy-D-glucose (FDG)
Lessor actuel en cancérologie de lutilisation in vivo , en
tomographie par émission de positons (TEP), du fluoro-
deoxyglucose (FDG) marqué au fluor 18 a réactualisé la
notion, établie depuis les travaux de Warburg (4,5), dune
hyperconsommation de glucose par glycolyse avec pro-
duction accrue de lactates, au détriment de la voie
oxydative, comme lune des principales caractéristiques
métaboliques des cellules cancéreuses. La possibilité dex-
ploiter cette augmentation de consommation de glucose
par les cellules tumorales pour détecter les tumeurs ou
mesurer leur activité métabolique est illustrée par de nom-
breuses études cliniques (6). Le FDG est un analogue du
2-déoxy-glucose qui, transporté à lintérieur de la cellule
par diffusion facilitée comme le glucose, est ensuite
phosphorylé en 6 par lhexokinase en fluoro-2-DG-6-phos-
phate. Celui-ci, nétant ni métabolisé ni déphosphorylé,
est trappé dans la cellule et sy accumule, permettant ainsi
la visualisation du tissu tumoral par TEP. Les premiers tra-
vaux qui ont établi laccumulation du FDG dans les tu-
meurs remontent aux années 80 sur des modèles animaux
(7-11). Les premières applications cliniques ont concerné
les tumeurs cérébrales (gliomes) (12).
BIOLOGIE DU FDG ET DES AUTRES TRACEURS
ÉMETTEURS DE POSITONS
UTILISÉS EN CANCÉROLOGIE
J.Ph. VUILLEZ
Biophysique et Médecine Nucléaire - Laboratoire dEtude de Radiopharmaceutiques (L.E.R.)
UPRESA CNRS 5077 - LRC CEA 10 - CHU et Faculté de Médecine
- Grenoble -

Biologie du FDG et des autres traceurs émetteurs de positons utilisés en cancérologie
5
Revue de l'ACOMEN, 1998, vol.4, n°1
Laugmentation de la glycolyse dans les cellules tumora-
les est sous-tendue par des modifications enzymatiques
(13,14), et par une augmentation du transport trans-membra-
naire et de la captation cellulaire du glucose. Ces deux
phénomènes, dont la part relative varie dune tumeur à
lautre. ne sont ni la traduction dun dérèglement "anar-
chique" du métabolisme tumoral, ni simplement réaction-
nels aux conditions microenvironnementales difficiles des
tumeurs (défaut de vascularisation, hypoxie). Ils résul-
tent dune programmation génétique parfaitement ordon-
née, sous la dépendance directe de la transformation et de
la progression tumorales. Cette programmation reproduit
dans une version pathologique (parce que permanente et
non régulée) les mécanismes physiologiques dadaptation
métabolique mis en jeu lors de la prolifération des tissus
sains : croissance, cicatrisation, renouvellement tissulaire.
Il ny a pas de différence fondamentale de la nature des
phénomènes entre les cellules proliférantes et les cellules
cancéreuses (15).
Les modifications du métabolisme glucidique dans les tu-
meurs sont donc liées à la transformation maligne. Elles ne
sont pas uniquement dépendantes de la vitesse de crois-
sance (16) mais interviennent en amont des phénomènes
de prolifération quelles contribuent à permettre et à favo-
riser. Elles ne sont pas secondaires aux processus du cy-
cle cellulaire, mais partagent un déterminisme commun avec
la mise en cycle et la prolifération des cellules. Cest pour-
quoi les oncogènes peuvent influer sur la consommation
de glucose indépendamment de leur effet sur le cycle cel-
lulaire (17).
Ces modifications ne sont pas obligatoires, mais confére-
raient un avantage sélectif aux cellules tumorales (18). La
mauvaise qualité du micro environnement, sans être le fac-
teur causal, intervient comme pression de sélection des
cellules ayant une capacité glycolytique élevée. Le fait
que laugmentation du transport du glucose et de la gly-
colyse soit quasi-constante dans les cellules transformées
suggère que ces modifications, même sans être indispen-
sables, reflètent des altérations sous-jacentes de la régu-
lation cellulaire essentielles dans le processus de trans-
formation (19).
1. Modifications du métabolisme
glucidique dans les tumeurs
Laugmentation de la glycolyse et laugmentation de la
captation cellulaire de glucose peuvent être perturbés de
façon dissociée (16). Ces altérations réorientent le méta-
bolisme cellulaire vers la synthèse des précurseurs des
acides nucléiques indispensable à la multiplication des cel-
lules.
1.1. Augmentation de la glycolyse aérobie
Les premières observations, maintenant anciennes (4,5),
ont été affinées mais jamais remises en cause ; en revan-
che, la place attribuée à cette perturbation dans lhoméos-
tasie cellulaire a évolué.
1.1.1. Rappel sur la régulation de la glycolyse dans les
cellules au repos (20)
Voie glycolytique et production de lactates
Dans les cellules normales, le niveau et la direction du
métabolisme glucidique résultent de ceux des deux voies
opposées de la glycolyse et de la glyconéogénèse, et dé-
pendent du contrôle de la quantité et de lactivité de cer-
taines enzymes «clefs» ; celles-ci sont au nombre de trois
dans le sens de la glycolyse : hexo/glucokinase (HK/GK),
phosphofructokinase (PFK) et pyruvate kinase (PK), et de
quatre dans le sens de la glyconéogénèse : glucose-6-
phosphatase (G6P), fructose-1-6-diphosphatase (FDP),
phosphoénolpyruvate carboxykinase (PEPCK), et pyruvate
carboxylase (PC). Lactivité de ces enzymes-clefs est régu-
lée, dans des sens opposés, par des molécules signal.
Lorsque lapport en oxygène est suffisant (Figure 1), les
cellules adaptent leur production de pyruvate au niveau
de consommation de lacétyl-CoA et à leurs besoins en
énergie, principalement par laction inhibitrice de lATP
sur la PFK. Cette inhibition diminue la conversion du fruc-
tose-6-P en fructose 1,6-bi-P. A létat stationnaire, on ob-
serve un taux bas de fructose 1,6-bi-P, rapidement converti
en pyruvate par les étapes suivantes (dont la principale
est catalysée par la PK), et en fructose-6-P par la fructose
1-6 diphosphatase. Ce faible niveau de fructose 1,6-bi-P
est insuffisant pour empêcher linhibition de la PFK par
lATP. Secondairement le glucose-6-P, qui saccumule en
raison de linhibition de la PFK, inhibe sa propre synthèse
par lhexokinase. Au total, la production mitochondriale
dATP contrôle la glycolyse, essentiellement par linhibi-
tion de la PFK, et ceci est désigné sous le terme deffet
Pasteur (21).
En anaérobiose, ou en cas de manque relatif doxygène, la
production dATP dans la mitochondrie à partir dacétyl-
CoA est bloquée. Le niveau dATP diminue, linhibition
de la PFK est levée (et la fructose 1-6 diphosphatase est
bloquée par laugmentation de lAMP). Le fructose 1,6-bi-
P saccumule et renforce laction de la PFK. La baisse de
G-6-P qui en résulte augmente lactivité hexokinase. Lef-
fet synergique de ces différents phénomènes permet à la
cellule dutiliser au maximum la voie glycolytique pour
maintenir la production dATP lors de lanoxie. Afin que le
NADH puisse être produit à un niveau normal (par la
glycéraldéhyde 3-P déshydrogénase), il doit être oxydé en
NAD+. Cette oxydation ne pouvant plus avoir lieu dans la
mitochondrie, le NADH est recyclé lors de la transforma-

J.Ph. VUILLEZ
6Revue de l'ACOMEN, 1998, vol.4, n°1
tion du pyruvate en lactate ; ce dernier est ensuite excrété
par la cellule.
Voie ("shunt") des pentoses
Parallèlement à la voie glycolytique, le ribose-5-P est syn-
thétisé par la voie oxydative du shunt des pentoses phos-
phates, à partir du glucose 6-P, avec production concomi-
tante de NADPH destiné à des processus de biosynthèse
(des acides gras notamment), et de glutathion réduit (GSH)
pour les processus de détoxification. Le ribose 5-P est en
équilibre direct avec le fructose 6-P et le glycéraldéhyde 3-
P. Les deux enzymes de la voie oxydative des pentoses
phosphates sont inhibées dans les conditions physiolo-
giques par lATP. La direction du shunt des pentoses est
déterminée essentiellement par la vitesse de conversion
du glycéraldéhyde 3-P en pyruvate. Lorsque celle-ci est
élevée, comme cest le cas en normoxie dans les cellules
normales, le ribose 5-P est rapidement métabolisé en fruc-
tose 6-P, glycéraldéhyde 3-P et finalement en pyruvate.
Ces phénomènes ont lieu dans la mesure où la pyruvate
kinase est maintenue active , cest à dire sous forme non
dimérisée et non phosphorylée; ceci résulte à la fois de ce
que les protéine-kinases sont inactives et de ce que
lisoenzyme de la pyruvate kinase exprimée dans les cellu-
les normales (type musculaire M1), nest pas un substrat
adéquat pour ces protéine kinases.
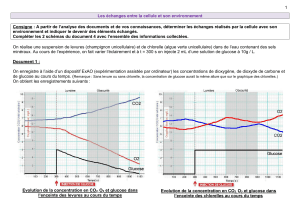
- Figure 1 -
Métabolisme glucidique et voie des pentoses phosphate dans les cellules normales.
Le point essentiel est linhibition de la phosphofructokinase par lATP générée dans la mitochondrie, expliquant linhibition de la
glycolyse en présence doxygène, cest-à-dire dans les conditions physiologiques (effet Pasteur).
(G-6-P = glucose-6-phosphate ; PFK = phosphofructokinase ; LDH = lactate deshydrogénase)
Le ribose-5-P est produit à partir du glucose 6-P, avec production concomitante de NADPH et de glutathion réduit (GSH). Le ribose 5-
P est en équilibre direct avec le fructose 6-P et le glycéraldéhyde 3-P. Les deux enzymes de la voie oxydative des pentoses phosphates
sont inhibées dans les conditions physiologiques par lATP. La direction du shunt des pentoses est déterminée essentiellement par la
vitesse de conversion du glycéraldéhyde 3-P en pyruvate. Lorsque celle-ci est élevée, comme cest le cas en normoxie dans les cellules
normales, le ribose 5-P est rapidemment métabolisé en fructose 6-P, glycéraldéhyde 3-P et finalement en pyruvate.

Biologie du FDG et des autres traceurs émetteurs de positons utilisés en cancérologie
7
Revue de l'ACOMEN, 1998, vol.4, n°1
1.1.2. Régulation de la glycolyse lors de la prolifération
cellulaire et dans les cellules tumorales
Les phénomènes physiologiques mettant en jeu la prolifé-
ration cellulaire comme la cicatrisation (22) ou la transfor-
mation lymphocytaire (23,24) saccompagnent de modifi-
cations du métabolisme glucidique qui sont retrouvées,
très exagérées, dans les cellules tumorales.
Dans les cellules qui prolifèrent, le blocage relatif de la
pyruvate kinase (qui catalyse la dernière étape irréversible
de la glycolyse) augmente la concentration de glycéraldé-
hyde 3-phosphate et favorise fortement laccumulation de
ribose-5-phosphate par la voie non oxydative des pento-
ses phosphate. Les taux élevés de fructose 1,6-diphos-
phate, également rencontrés dans ces conditions, inhibent
la 6-phosphogluconate déshydrogénase et réorientent la
voie des pentoses dans sa direction non oxydative. La
concentration de ribose-5-phosphate augmente. La syn-
thèse de P-ribose-PP est augmentée et la capacité de pro-
duction du NADPH et du GSH est réduite. Parallèlement
on observe une perte de leffet Pasteur et un niveau élevé
de glycolyse en conditions aérobies (21,23-25). Laugmen-
tation de la glycolyse semble liée à une reprogrammation
cellulaire, résultant de modifications enzymatiques sac-
compagnant dune inhibition de la voie oxydative (effet
"Pasteur inversé" ou effet Crabtree (21,26).
1.1.2.1. Modifications enzymatiques
Diverses modifications des enzymes de la glycolyse ont
été mises en évidence, à la fois quantitatives et qualitati-
ves (expression disoenzymes différents, avec pour corol-
laire changement des paramètres cinétiques). Dans
lhépatome de Novikoff se produit un changement
disoenzyme, avec le remplacement du type "hépatique"
de Km élevé sensible aux régulations nutritionnelle et hor-
monale, par des isoenzymes de type "non-hépatique", de
Km bas et ne répondant pas ou peu aux mécanismes de
régulation et de rétrocontrôle (27). Des revues détaillées
ont été publiées (13,14,18,28). Les phénomènes les plus
importants concernent les trois enzymes clefs que sont
lhexokinase, la phosphofructokinase et la pyruvate kinase.
Dans les cellules tumorales, la "reprogrammation généti-
que" de ces enzymes-clefs aboutit à un nouvel équilibre
par rapport à la cellule normale, qui favorise la glycolyse
au détriment de la néoglycogenèse.
Modifications de lhexokinase
Knox et al. (29) dans un modèle dhépatome (hépatome de
Morris) avaient déjà trouvé un taux dhexokinase par
gramme de tissu plus élevé dans les tumeurs que dans le
foie normal ; surtout, ce taux était corrélé à la vitesse de
croissance de la tumeur. Dans ce même travail les auteurs
soulignaient que le rendement de cette augmentation de
lhexokinase (HK) dans les hépatomes est médiocre, puis-
que le foie normal en régénération présente une vitesse de
croissance similaire avec un contenu en hexokinase envi-
ron deux fois moindre.
Laugmentation de lactivité HK a par la suite été large-
ment confirmée (30-36). Dans les tumeurs, 50 à 80 % de
lactivité HK est préférentiellement liée à la membrane ex-
terne des mitochondries, ce qui favoriserait lhypercatabo-
lisme du glucose (28,32,37,38). En effet, lassociation ré-
versible de lHK aux mitochondries est un mécanisme im-
portant de régulation de lactivité de cette enzyme : la forme
liée nétant pas (ou très peu) sensible à linhibition par le
glucose-6-phosphate comparativement à la forme soluble,
et pouvant utiliser directement lATP généré dans la mito-
chondrie (33), représente la forme la plus active (36). Ceci
est lié à lexpression préférentielle des isoenzymes I et II,
différents de ceux rencontrés dans les cellules normales ;
cest du moins à ces formes que sapparente lhexokinase
liée aux mitochondries (35). Il existe une relation entre le
degré de transformation maligne et laltération progres-
sive de la proportion des isoenzymes et de leur distribu-
tion subcellulaire (36). Le gène dune hexokinase tumorale
liée aux mitochondries a été cloné et séquencé (39).
Lactivité HK a été mesurée dans des cellules tumorales à
des niveaux similaires à celui de cellules normales, incom-
patibles avec laugmentation de la glycolyse aérobie cons-
tatée dans les tumeurs, conduisant à découvrir, dans tou-
tes les lignées tumorales testées, une importante activité
de type glucokinase (hexokinase de type 4 à Km élevé),
normalement rencontrée dans le foie et qui peut donc être
exprimée dans les cellules tumorales (25,40).
Modifications de la phosphofructokinase (PFK)
Moins de publications ont été consacrées à ce deuxième
enzyme clef de la glycolyse, dont le rôle est cependant
primordial. Il existe une augmentation de lactivité PFK
dans les tumeurs hépatiques, variable en fonction du de-
gré de différenciation (41,42,43), et accompagnée dune
modification du profil dexpression des isoenzymes (43,44).
Il n apparait pas de formes spécifiques des tumeurs, mais
les isoenzymes rencontrés dans les tissus normaux sont
exprimés en proportions anormales, dans des
glioblastomes et des astrocytomes) (45).ou dans des li-
gnées de leucémies et de lymphomes (44). Les différentes
formes immunologiques et électrophorétiques de lenzyme
résultent de sa structure tétramérique qui associe de façon
variable trois types de sous-unités (M, L ou P) codées par
trois gènes différents (44). La PFK est régulée par diffé-
rents métabolites, hormones et facteurs nutritionnels ou
liés au développement. Elle joue un rôle essentiel dans le
métabolisme énergétique, particulièrement dans les tissus
où la glycolyse est prédominante voire exclusive (érythro-
cytes matures, muscle lors de lexercice, cerveau, cur
ischémique) et de la même façon dans les tumeurs. Leffet

J.Ph. VUILLEZ
8Revue de l'ACOMEN, 1998, vol.4, n°1
Pasteur est expliqué par linhibition de la PFK par lATP, le
citrate, et dautres métabolites (21). Lexpression préféren-
tielle dans les tumeurs de lisoenzyme hépatique, qui pos-
sède une cinétique et une régulation particulières et nest
pas sensible à ces métabolites, explique au moins en partie
la perte de leffet Pasteur (44). De plus laccumulation de
fructose 1,6-biphosphate résultant de laugmentation de
lactivité HK entraîne une activation de la PFK qui peut
prendre le pas sur linhibition par lATP (20). Ce
rétrocontrôle positif est favorisé par lexpression disoenzy-
mes particulièrement sensibles au fructose 1,6-biphosphate
(44,46). Il en résulte secondairement un "emballement" de
la PFK qui sajoute à laugmentation de lactivité HK.
Modifications de la pyruvate kinase
Lactivité de la PK est très élevée dans les cellules tumora-
les (25,28,31). Dans des tumeurs du sein, cette augmenta-
tion de lactivité PK est corrélée au degré de malignité par
référence au tissu mammaire sain (47). Parallèlement, on
assiste à lexpression dune forme modifiée, caractéristi-
que des cellules tumorales, de la PK (48) qui a été bien
montrée dans les tumeurs du sein (47) et retrouvée dans
les tumeurs cérébrales (49) et pratiquement toutes les tu-
meurs et les cellules proliférantes (28).
Lisoenzyme de type tumoral PK-M2 est soumis à une ré-
gulation particulière et complexe par divers métabolites :
inhibition par des acides aminés (alanine, phénylalanine)
(48,49), activation par le fructose 1,6-biphosphate (48) ;
dautres facteurs sont discutés mais apparaissent moins
importants (48) ; la PK-M2 est également inhibée par lATP
(48). Son affinité pour le phosphoénolpyruvate est plus
élevée que celle de lisoenzyme L rencontrée dans le foie,
mais plus basse que celle de lisoenzyme M1 (PK-M1) ren-
contrée dans le muscle (50). La régulation de la PK-M2 fait
par ailleurs intervenir un phénomène de phosphorylation
hormono et oncogène-dépendant (28,51). La PK-M2 peut
en effet être phosphorylée (à la différence de la PK-M1) par
une protéine kinase AMPc-indépendante ainsi que par la
tyrosine kinase codée par loncogène PP60v-src. Cette
phosphorylation inactive lenzyme. La protéine kinase
AMPc-indépendante se lie avec une affinité très élevée à
la PK-M2 et en catalyse la phosphorylation, préférentielle-
ment lorsque celle-ci est sous forme dimère.
L activité de la PK-M2 présente ainsi une très grande va-
riabilité (28) : elle peut être extrêmement élevée ou au con-
traire, (comme celle de la PK normale à certains moments
du cycle cellulaire), être virtuellement égale à zéro. Cette
inactivation, qui survient en réponse à la régulation exer-
cée par des oncogènes ou des facteurs de croissance, ex-
plique quil existe une bonne corrélation entre lactivité
PK maximale et le degré de malignité, mais pas avec le
niveau de glycolyse ni la production de lactates. Au total
on assiste dans les cellules tumorales à un blocage relatif
de la PK-M2.
Autres enzymes
Des modifications portant sur les autres enzymes de la
glycolyse ont été décrites mais leur rapport avec laug-
mentation de la glycolyse est moins direct. On en trouve
des études détaillées dans la littérature (25,31).
Mécanismes des modifications enzymatiques
Les différentes modifications enzymatiques envisagées,
aussi bien quantitatives que qualitatives, sexpliquent avant
tout par des régulations au niveau du génome. Ceci est
bien documenté pour lhexokinase, dont laugmentation
dactivité est largement dépendante de lactivation de la
transcription (52,53). Rempel et al. (54) ont montré une am-
plification du gène de lhexokinase de type II (HK II) dans
une lignée cellulaire dhépatome de rat. Pour ces mêmes
auteurs (38), la transcription du gène dans les tumeurs est
indépendante de létat métabolique, conférant un avan-
tage sélectif aux cellules cancéreuses. Le promoteur de
lHK II est sous le contrôle du glucose lui-même (55). Des
mécanismes post-transcriptionnels et même post-
traductionnels peuvent être impliqués (56), notamment la
phosphorylation de certaines enzymes.
1.1.3.2. Modifications des mitochondries et altérations
de la respiration cellulaire
La responsabilité dune anomalie primitive des mitochon-
dries, corollaire dune défaillance de la respiration cellu-
laire, dans laugmentation de la glycolyse, était la théorie
initialement avancée par Warburg. Même si cette théorie
est maintenant récusée, il demeure que les mitochondries
présentent, dans les tumeurs, des anomalies dont on con-
naît mal la signification.
Les observations réalisées grâce aux cations lipophiles
comme le tétraphénylphosphonium et surtout la rhodamine
123, en présence de divers ionophores, ont permis de met-
tre en évidence une augmentation très sensible de la ddp
mitochondriale (60 mV environ) dans les cellules tumora-
les (57). On sait que ces données ont été confirmées par
les travaux de Piwnica-Worms avec le 2-méthoxyisobutyl
isonitrile (MIBI) dont la rétention cellulaire est liée, comme
pour la rhodamine 123, aux différences de potentiel trans-
membranaires (58,59). Lorigine de cet accroissement de la
ddp mitochondriale nest pas connue, elle est de plus in-
constante (57) : elle est induite par loncogène v-fos, alors
que loncogène v-fes a leffet inverse ; elle est très mar-
quée dans les tumeurs bien différenciées mais pratique-
ment non retrouvée dans les tumeurs peu différenciées
comme les lignées FET et CCL 237 de cancer colique hu-
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%