De la variabilité génétique à la sélection De la variabilité

Terminale S
Partie 3 – Stabilité et variabilité des génomes - Évolution
Fiche #12 – Les innovations génétiques
De la variabilité génétique à la s
é
lection
10 Les fréquences des allèles d’un gène, dans une population donnée, ne sont pas égales. Comment expliquer qu’un allèle
soit plus fréquent qu’un autre ?
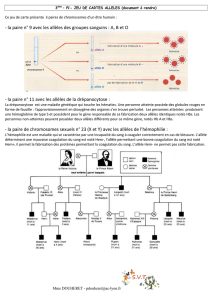
10.1 I/C Décrivez les différents phénotypes possibles pour la phalène du bouleau (doc. 3.3 et 148.A).
À quel milieu chaque phénotype semble-il adapté ?
10.2 Ra/C D’après les docs. 3.4 et 149.B2, rédigez les étapes de la démarche expérimentale adoptée par Kettlewell (rappel
des étapes : Observation, Problème, Hypothèse, protocole Expérimental, Résultats, Interprétation, Conclusion).
10.3 Re/C En utilisant le tableur Excel, représentez sous forme d’un graphique l’évolution des captures de phalènes
(onglet ‘Exp. De Kettlewell’), ainsi que l’évolution de la fréquence des formes typica et carbonaria au cours des 100
dernières années (onglet ‘Fréquences allèles phalène’)
10.4 I/Re À l’aide du logiciel de simulation Sélection naturelle (http://darwin.eeb.uconn.edu/simulations/selection.html),
testez l’influence de l’avantage sélectif apporté par une mutation sur sa fréquence dans une population, avec :
p = 0,1 fréquence de départ de l’allèle C
w11 = 1,0 avantage sélectif d’un individu homozygote (C//C)[C] dans un milieu pollué
w12 = 1,0 avantage sélectif d’un individu hétérozygote (C//c)[C] dans un milieu pollué
w22 = 0,6 (dés)avantage sélectif d’un individu homozygote (c//c)[c] dans un milieu pollué
Faites varier w12, en envisageant le cas d’une codominance. Interprétez la courbe obtenue.
Puis modélisez le cas d’une population de type carbonaria dont l’environnement devient non pollué.
10.5 C Tracez les résultats obtenus sur un graphique
De la variabilité génétique sans sé
lection
11 Certaines mutations, à cause du caractère dégénéré du code génétique, n’ont aucune répercussion sur le phénotype
(mutations silencieuses). Comment ces mutations se propagent-elles dans une population ?
11.1 I/Ra Interprétez les résultats du doc. 153.B3
11.2 I/Re À l’aide du logiciel de simulation Dérive génique (http://darwin.eeb.uconn.edu/simulations/drift.html), testez
l’influence de la taille de la population sur l’évolution de la fréquence d’une mutation silencieuse. Résumez les résultats
obtenus, en les comparants au doc. 3.5.
Maintien d’un désavantage sélectif
12 Comment un allèle constituant un désavantage sélectif peut-il se maintenir dans une population ?
12.1 I/C Comparez les aires de répartition du paludisme et de la drépanocytose (doc. 150.A)
12.2 I/C Commentez le document 3.6.
12.3 Ra Mettez en relation les deux informations précédentes.
12.4 Ra/Re Trouvez les valeurs appropriées pour simuler ce cas à l’aide de simulation Sélection naturelle.

Terminale S
Partie 3 – Stabilité et variabilité des génomes - Évolution
Fiche #12 – Les innovations génétiques
Document 3.3 – Fréquence
des phénotypes carbonaria et
typica de la phalène du
bouleau et localisation des
régions industrielles (en
foncé). Dans les régions
industrielles, la pollution est
responsable de la mort des
lichens. Données de 1950.
Les observations En Grande-Bretagne, la
forme noire (mélanique) [de la phalène du
bouleau] fut signalée pour la première fois en
1848. Par des croisements contrôlés, le
biologiste anglais Kettlewell et son équipe ont
montré dans les années 1950 que le
phénotype mélanique est dû à un allèle
dominant (C) et que les individus clairs sont
homozygotes pour l'allèle récessif (c).
À partir de la fréquence des phénotypes typica
et carbonaria, les généticiens ont calculé les
fréquences des deux allèles dans la population
de phalène de la région de Manchester entre
1848 et 1948. Cette fréquence est le rapport,
au sein d'une population, du nombre
d'exemplaires de l'allèle considéré au nombre
total d'exemplaires des allèles pour ce gène.
C
c
1848
0.00 1.00
1858
0.00 1.00
1868
0.03 0.97
1878
0.45 0.55
1888
0.76 0.24
1898
0.86 0.14
1908
0.90 0.10
1918
0.92 0.08
1928
0.94 0.06
1938
0.96 0.04
1948
0.96 0.04
Fréquence allèlique
Document 3.4 – Expériences
de Kettlewell et leur
interprétation théorique.
Dans les années 1950, le biologiste Kettlewell et son
équipe ont noté le nombre d'individus des deux
formes capturés par piégeage lumineux dans une
région industrielle (Birmingham) et dans une région
rurale (la forêt de Dean End Wood) (expérience 1).
Ils lâchèrent ensuite des papillons marqués des
deux formes dans chacune des deux régions
(expérience 2). Quelques jours après, ils ont
déterminé le nombre d'individus marqués, des deux
formes, recapturés dans les deux sites. Enfin, ils ont
exposé à la prédation par les oiseaux le même
nombre de papillons des deux formes sur chacun
des deux sites (expérience 3) et ils ont mesuré le
nombre de papillons capturés dans chaque cas.
« Pour qu'il y ait sélection naturelle, quatre
conditions doivent être remplies. [ ...] Il faut
que les organismes se reproduisent pour
engendrer une nouvelle génération [...] que
cette reproduction soit globalement conforme
[c'est-à-dire qu'il y ait hérédité]. [...] Les
caractères individuels héréditaires doivent
varier parmi les membres d'une population. [...]
[Il faut] qu'un certain état d'un caractère
héréditaire confère, [dans certaines conditions
de l'environnement], à certains membres de la
population une probabilité de reproduction plus
grande que les autres [c'est-à-dire un
avantage sélectif].»
Marc Ridley, Évolution Biologique, De Boeck,
1997, pp. 71-72
Document 3.5 – Vitesse
d’évolution de différents
gènes
Protéine ou
peptide
Fonction de la protéine ou du peptide Vitesse
d'évolution
Fibrinopeptide Fragment du fibrinogène. Son excision permet la transformation du
fibrinogène en fibrine, protéine qui intervient dans la coagulation du
sang. Le fibrinopeptide libéré n'a pas de fonction connue. 8,3
Ribonucléase
pancréatique
Enzyme digestive 2,1
Globine α Transport du dioxygène dans les érythrocytes 1,2
Cytochrome C Molécule de la mitochondrie indispensable à la respiration
cellulaire dans toutes les cellules.
0,3
À partir de la date de divergence entre deux lignées de deux espèces (déterminée grâce à la paléontologie) et d'une
comparaison de la séquence d'une protéine entre ces deux espèces, on peut calculer une vitesse d'évolution globale
de la protéine. Elle s'exprime en nombre de substitutions par acide aminé et par 109 ans. Il est de même possible de
comparer la vitesse d'évolution de différentes régions d'une même protéine.
Document 3.6 – Prévalence de
l’allèle drépatnocytaire S
(gauche) et du paludisme
(droite)
Impossible d'afficher l'image liée. Le fichier a peut-être été déplacé, renommé ou supprimé. Vérifiez que la liaison
pointe vers le fichier et l'emplacement corrects.
Impossible d'afficher l'image liée. Le fichier a peut-être été déplacé, renommé ou supprimé. Vérifiez que la liaison
pointe vers le fichier et l'emplacement corrects.
1
/
2
100%