I_genetique_et_evolution 2127KB Sep 18 2013 05:59:40 PM

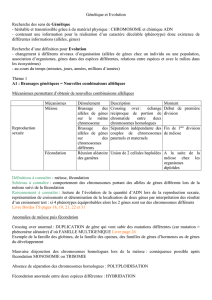
Chapitre I Génétique et évolution.
Problème: Comment expliquer la biodiversité actuelle ?
Définition: La biodiversité est la diversité naturelle des organismes vivants. Elle s'apprécie en
considérant la diversité des écosystèmes, des espèces, des populations et celle des gènes.
A Le brassage génétique et la diversité des génomes.
II Reproduction sexuée et stabilité du caryotype.
Comment les mécanismes de la reproduction sexuée assurent-ils la stabilité d'une espèce?
Bilan
Au cours du cycle de développement d'une espèce animale à reproduction sexuée, il y a alternance d'une
phase diploïde, et d'une phase haploïde. Cependant les organismes sont soit majoritairement diploïdes ou
haploïdes. L Homme est donc une espèce diploïde de formule chromosomique 2n=46, c 'est à dire que l'on
trouve 23 paires de chromosomes homologues dans les cellules somatiques et 23 chromosomes dans les cellules
sexuelles.
Cette stabilité du caryotype au fil des générations est assurée par l'alternance, au cours du cycle de
développement, de deux processus biologiques complémentaires: la méiose qui permet de passer de la phase
diploïde à la phase haploïde, alors que la fécondation permet le passage de la phase haploïde à la phase diploïde.
La fécondation correspond à la fusion d'un spermatozoïde et d'un ovule (ovocyte pour l'Homme) formant ainsi
une cellule œuf (zygote), première cellule d'un nouvel individu.
La méiose étant une phase de la production des gamètes (cellules sexuelles: spermatozoïdes ovocytes) par les
gonades (testicules ou ovaires).
( Il existe aussi des organismes haploïde comme Sordaria qui est une espèce de champignon de formule
chromosomique n=7)

Schéma d'un chromosome à 2 chromatides.
III Le déroulement de la méiose.
Comment la méiose permet-elle le passage de cellules diploïdes à des cellules haploïdes ?
Bilan:
La méiose est un processus biologique qui permet le passage d'une cellule mère diploïde à 4 cellules filles
haploïdes . Elle est constituée d'une succession de deux divisions cellulaires et elle est précédée d'une phase de
réplication de l'ADN dans l'interphase (double le matériel génétique).
Au cours de la première division de méiose (division réductionnelle) on retrouve 4 étapes.
La première étape est la prophase I, il se forme les chromosomes, la membrane nucléaire disparaît et il y a
appariement des chromosomes homologues (forme des paires qui se nome bivalent ou tétrade) pour la cellule
mère.
La métaphase I les bivalents (paires de chromosomes homologues) migrent à l'équateur de la cellule.
L'anaphase I Séparation des deux chromosomes de chaque paire, qui migrent aux pôles opposés de la cellule.
La télophase I et prophase II, séparation du cytoplasme en 2 et création de 2 cellules filles. La prophase II est le
début de la deuxième division (division équationnelle).

La métaphase II Les chromosomes se placent à l'équateur de la cellule.
L'anaphase II Les chromosomes se séparent en deux chromosomes à une chromatine et chaque chromatine
migre à un pôle de la cellule.
La télophase II, séparation du cytoplasme en 2 et création de 4 cellules filles haploïdes.
http://www.prepaangers.fr/cariboost1/crbst_181.html
IV Le brassage intrachromosomique (le crossing-over)
Bilan
Le phénotype, correspond à l'ensemble des caractères étudiés pour un individu, un groupe ou un espèce,
pour un caractère donné on trouve plusieurs phénotypes possibles, le phénotype le plus couramment observé dans
la nature est appelé « sauvage », les autres sont en général des phénotypes mutants.
Ce phénotype est sous la dépendance d'un ou plusieurs gènes, eux même pouvant avoir un ou plusieurs allèles.
Le génotype correspond donc à la combinaison des gènes et des allèles responsables du caractère étudié. Certains
individus sont porteurs d'allèles identiques pour certains gènes, ces individus sont dit homozygotes (souche ou
lignée pure), dans le cas d'individus porteurs de deux allèles différents pour un même gène ils sont alors
hétérozygotes.

Exemple: caractère étudié groupe sanguin, pour un individu de groupe sanguin A son phénotype sera (A) et son
génotype peut être [A//o] car l'allèle A est dominant et donc écrit en majuscule et l'allèle o est en minuscule car il
est récessif. Si les deux allèles sont dominants c’est le cas du phénotype (AB) on parle de codominance des
allèles. Dans les cas étudiés on étudie généralement des cas ou il y a un seul gène responsable d'un caractère.
Pendant la prophase I, les chromosomes homologues se regroupent (sous forme de tétrade ou bivalents), et
entrecroisent leurs chromatides: on parle d'appariement. Au niveau des chiasmas des portions de chromatides des
2 chromosomes peuvent s'échanger, il y a alors formation de chromosomes remaniés. Des combinaisons non
parentales d'allèles apparaissent sur une chromatide, ce processus est appelé crossing-over. Il y a création de
gamètes recombinés et donc après fécondation formation de phénotypes recombinés. Dans le cas de croisements
tests, le pourcentage de phénotypes recombinés est plus faible que le pourcentage de phénotypes parentaux car
ce phénomène de crossing-over est en général peu fréquent.
Photo de bivalent.

Crossing-over
Phénomène génétique de la méiose où les deux chromosomes de chaque paire se rapprochent et se brisent au
même niveau de la séquence génétique pour ensuite échanger les fragments résultants.
Chiasmas
Points de chevauchement de deux chromatides, observables en microscopie.
Le test-cross ou croisement test a pour but de révéler le génotype d'un organisme qui présente un phénotype
dominant. Cet individu peut être soit hétérozygote, soit homozygote pour l'allèle dominant. Le moyen le plus
efficace de connaître son génotype est de le croiser avec un organisme exprimant le phénotype récessif. Les
phénotypes de la génération suivante permettront de déterminer le génotype du parent ayant un phénotype
dominant.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%