Neuroplasticité : Aspects Développementaux, Corticaux et Synaptiques

1
LA NEUROPLASTICITE
G. GANDOLFO
Laboratoire de neurobiologie et psychotraumatologie
Université de Nice-Sophia Antipolis
Introduction
Obsolescence de la controverse inné-acquis
La plasticité développementale
Notion de période critique : le chant des oiseaux
Influence du milieu extérieur : le développement du cortex visuel
Influence du milieu intérieur : la différenciation sexuelle chez le fœtus de mammifère
La plasticité comportementale adulte (ou postdéveloppementale)
La vicariance
La suppléance
L’adaptation motrice chez le spationaute en apesanteur
La malléabilité des perceptions affectives
La plasticité corticale
La neurogenèse secondaire
Les cartes corticales sensorielles et motrices
La plasticité synaptique
Notion d’assemblée cellulaire et de force (ou efficacité) synaptique
L’apprentissage par stabilisation sélective des synapses en voie de développement
Plasticité à court et à long terme
Notion de métaplasticité
La plasticité réparatrice
La capacité d’autoréparation du système nerveux :
La régénération dans le système nerveux périphérique
La régénération centrale : bourgeonnement, hypersensibilité de dénervation, synapses
silencieuses
La médecine régénérative : thérapies cellulaires, rééducation ciblée, stimulation intracérébrale
Conclusion
Les enjeux éthiques de la recherche biomédicale sur la neuroplasticité
Pour en savoir plus :
Gandolfo, G. et Grammont, F. Les divers aspects de la neuroplasticité. Biologie-Géologie,
2 : 291-312, 2005.
Gandolfo, G. et Miquel, P.A. La mémoire : une approche interdisciplinaire. Biologie-
Géologie, 2 : 97-130, 2008.

2
Obsolescence de la controverse inné-acquis
Longtemps on a pensé que le SN était intangible, immuable : dès sa constitution, le
cerveau était une structure organique fixée une fois pour toutes et commençait à mourir dès
l’adolescence. Autrement dit, l’aspect inné était prépondérant.
Les progrès techniques réalisés en génétique ont permis de décrypter à tour de bras
surtout à partir des années 2000 les génomes de nombreux êtres vivants. On s’est rendu
compte qu’il n’y avait pas de grosses variations entre Caenorhabditis elegans, un vers
nématode, et le chimpanzé : de 20 à 35 000 gènes. Puis ce fut le tour du génome humain en
2009 : une petite trentaine de milliers de gènes seulement, un chiffre revu récemment à la
baisse d’ailleurs (22 à 25 000 selon les études), bref, comme la plupart des animaux. Il est loin
le temps où l’on s’était crédité d’au moins 100 000 gènes… avant toute technique de
décryptage ! Pire si l’on considère certains végétaux : la tomate a aussi 35 000 gènes et le
maïs en possède 54 600 soit plus du double que l’Homme! En dépit de la blessure infligée à
notre orgueil anthropocentriste, il a fallu convenir que ce nombre de gènes est largement
insuffisant pour expliquer l’immense variété des caractères humains, de nos conduites
comportementales, de nos façons de faire, d’agir, de savoir, car il y a autant de personnalités
que de personnes vivant et ayant vécu sur Terre. Impossible donc d’expliquer avec ce nombre
de gènes le fonctionnement du cerveau et du SN, un système aussi dynamique, aussi
redondant, qui est capable de s’adapter aux innombrables variations des milieux (extérieur et
intérieur) et d’être en fin de compte modulé par la propre activité qu’il génère.
Qu’est-ce qui fait alors la différence ? Alors que le nématode C. elegans possède 302
neurones et la mouche du vinaigre 200 000, on admet aujourd’hui que le cerveau humain est
constitué d’environ 100 milliards de neurones, chacun étant capable d’établir en moyenne un
millier de connexions synaptiques avec les autres (réseaux), ce qui représente quelques cent
mille milliards de synapses ! De quoi expliquer les très nombreuses aptitudes du cerveau et
surtout son extraordinaire faculté d’adaptation, qu’on appelle la plasticité. C’est donc bien les
incessantes réorganisations de ces réseaux de neurones, et non pas le nombre de gènes, qui
sont à l’origine de l’extrême variabilité des capacités humaines. Ex : qu’est-ce qui fait qu’un
pianiste est moyen, bon ou virtuose : ce ne sont pas ses gènes, mais le fonctionnement plus ou
moins coordonné de ses réseaux de neurones.
Certes, l’influence des gènes demeure : ils donnent un cadre dans la constitution du
SN mais aussi dans le changement de la connectivité neuronale, autrement dit ils rendent
possibles les modifications de certains comportements. Comme le prouvent les animaux ayant

3
subi des mutations génétiques expérimentales : souris avec des troubles de la locomotion, de
la capacité d’apprentissage, de la prise alimentaire ou hydrique… (modèles animaux).
Mais si les gènes donnent un cadre à la construction et au fonctionnement du cerveau,
les réseaux de neurones aussi, qui sont constamment modifiés par les facteurs
environnementaux liés à l’expérience du sujet (apprentissage). Quel est alors leurs poids
respectif, entre déterminisme génétique et liberté personnelle ? Notre sentiment de liberté
dans nos prises de décisions morales (libre-arbitre) est-il réel ou illusoire ? D’autant qu’on vit
en société. Certes, certaines sociétés (insectes sociaux) sont organisées de façon entièrement
génétique avec une seule reine (et pas 2 !), des ouvrières, des soldats, chacun ayant une tâche
bien précise. Mais si, par ex, les butineuses chez les abeilles sont décimées, le programme
génétique fera alors en sorte que des ouvrières vont donner des butineuses.
Au cours de l’évolution des espèces, les gènes vont perdre progressivement le contrôle
final des comportements. Ex : chez la mouche du vinaigre, le programme génétique contrôle
encore entièrement le comportement sexuel. Mâles et femelles sont normalement
hétérosexuels à 100% et le mâle suit un protocole de séduction rigide : faire de la musique
avec ses ailes, toucher les pattes de la femelle qui a émis une phéromone… Si, par
manipulation génétique, on empêche la femelle de produire les phéromones, le mâle va alors
courtiser… d’autres mâles ! Mâle et femelle ont un gène commun « responsable » de ce
comportement mais qui est « travaillé » (épissé) de façon différente selon le sexe : si on
transfère ce gène d’un mâle chez une femelle, celle-ci va courtiser d’autres femelles, comme
l’aurait fait le mâle.
A ne pas extrapoler à l’Homme ! Notre constitution génétique est faite pour qu’on soit
libre : si les réseaux de neurones se construisent bien à partir du programme génétique
(« circuiterie » neuronale), les détails de la construction dépendent de l’environnement. Ex. du
morceau de musique : il y a la partition d’origine écrite par le compositeur et les variations
jouées par le pianiste qui vont changer la mélodie, chaque pianiste apportant sa « touche »
personnelle.
Ce qui change, c’est la force (efficacité) des connexions synaptiques : la plasticité tend
à nous rendre tous différents, même des jumeaux avec certains caractères communs mais qui
ont des personnalités différentes, une différence encore plus marquée s’ils ont été élevés dans
des milieux différents (adoption séparée après l’orphelinat). Les réseaux de neurones et les
connexions synaptiques qui composent le SN sont ainsi constamment modifiés par
l’apprentissage, même chez l’adulte. Ces modifications peuvent même être transmises de
génération en génération en dehors de la voie des gènes : on parle d’épigenèse et de

4
mémétique. L’épigenèse (ou épigénétique) est l’ensemble des facteurs environnementaux
(nutritionnels, sensoriels, l’expérience sociale) et intrinsèques (interactions entre les cellules,
sécrétions de substances chimiques : neuromédiateurs, hormones…) qui sont à la base surtout
de la plasticité développementale. La mémétique est l’étude de la transmission dans le temps
et l’espace des activités culturelles au sein des communautés humaines : le « même » est un
terme forgé par Richard Dawkins (Le gène égoïste, 1976) et désigne un réplicateur
indépendant de l’ADN et permettant de transmettre, en dehors donc de la voie génétique, une
unité d’information culturelle (idée, mode de pensée, savoir-faire, technique, habitudes,
traditions, us et coutumes…) qui se copie d’un cerveau vers un autre selon des processus
évolutionnistes ; ce sont surtout des apprentissages par imitation dans lesquels interviennent
les neurones miroirs. On est ainsi arrivé à ce paradoxe : les progrès dans les techniques
génétiques, dont le décryptage génomique, a mis un terme à l’idéologie du « tout génétique »
qui réduisait tout comportement, toute pathologie à l’expression d’un ou plusieurs gènes.
La neuroplasticité souligne donc le caractère obsolète du débat séculaire entre l’inné et
l’acquis. Charles Darwin fut le premier à le remarquer quand il dit que la sélection naturelle
ne peut se faire sans des variations spontanées de l’instinct : il définit l’adaptation comme un
dispositif permettant à la fois de saisir les occasions génétiques et de diriger le hasard vers des
voies compatibles avec la vie dans un milieu donné. L’empreinte perceptive, popularisée par
Konrad Lorenz, entérine cette obsolescence : la possibilité d’empreinte à la naissance qui se
traduit par un attachement profond et durable est innée car toutes les espèces évoluées
(certains poissons, les oiseaux et les mammifères la possèdent), mais l’attachement est une
acquisition dans la mesure où Lorenz s’est substitué à la mère naturelle des oisons. François
Jacob (Le jeu des possibles) fera, lui, de l’adaptation le résultat d’un dialogue permanent entre
les gènes et le milieu environnant, entre les facteurs biologiques et culturels.
La plasticité du SN s’organise ainsi à plusieurs niveaux : au cours du développement
de l’individu lors des premiers âges de la vie, mais aussi chez l’adulte. Elle concerne, de la
manière la plus visible, les comportements et leur malléabilité, mais également le niveau
structural du cerveau, notamment le cortex cérébral, siège des fonctions cognitives, qui a
l’aptitude à se modifier en fonction des diverses expériences vécues, et enfin le niveau
cellulaire et synaptique. D’où le plan (doc.1) : plasticité développementale (avec la notion de
période critique) ; plasticité comportementale chez l’adulte ; plasticité corticale ; plasticité
synaptique ; enfin, l’aspect clinique avec la plasticité réparatrice, c’est-à-dire la capacité
d’autoréparation du SN après lésions et les pistes thérapeutiques qu’elle promet dans la
médecine régénérative.

5
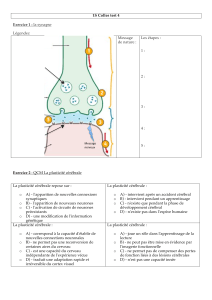
La plasticité développementale
1/ Notion de période critique (ou sensible) au cours de l’ontogenèse (doc.2)
Cas du chant des oiseaux dans diverses situations expérimentales montrant, selon les
espèces, des différences de plasticité de la structure cérébrale du chant.
2/ Influence du milieu extérieur : le développement du cortex visuel (doc.3 et 4)
Doc.3 : les expériences de Riesen chez le chaton reprises par Hubel et Wiesel (Prix
Nobel) montrent qu’une fonction donnée peut être abolie sans retour possible si elle n’est
jamais exercée durant la période critique : si on suture la paupière d’un œil (privation
monoculaire) avant l’âge de 2,5 mois après la naissance, le nombre de neurones dans le cortex
visuel primaire est drastiquement réduit et l’animal, devenu adulte et auquel on enlève la
suture, se comporte comme un animal borgne (il perd sa vision stéréoscopique). Ce n’est plus
le cas si la suture palpébrale est effectuée plus tard, même si le nombre de neurones reste
globalement diminué par rapport au témoin.
Doc.4 : même expérience faite sur le singe et montrant au moyen d’un traceur
radioactif l’altération de la mise en place des colonnes de dominance oculaire qui donnent son
aspect (et son nom) strié au cortex visuel.
3/ Influence du milieu intérieur : la différenciation sexuelle chez le fœtus de
mammifère ;
La période critique est, ici, intra-utérine. La différenciation sexuelle se fait en fonction
de la présence ou non de testostérone, l’hormone sexuelle mâle, le cerveau du fœtus prenant
alors les caractères sexuels correspondants. Tout se joue sur l’hypothalamus, une structure
cérébrale responsable des variations mensuelles de la sécrétion des hormones sexuelles. Si un
événement extérieur modifie l’ambiance hormonale lors de la période critique fœtale, on peut
modifier définitivement les caractères sexuels ultérieurs et provoquer certaines déviations.
Une étude sociologique a ainsi montré en Allemagne un fort accroissement du taux
d’homosexualité masculine chez les garçons nés vers la fin de la Seconde Guerre mondiale :
le stress maternel des femmes enceintes, en libérant massivement de l’ocytocine, a neutralisé
les effets de la testostérone.
En conclusion, l’environnement joue un rôle indispensable dans l’établissement d’une
aptitude, mais si les stimulus adéquats ne surviennent pas au cours d’une période critique (de
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
6
7
8
9
10
11
12
13
14
15
16
17
18
19
1
/
19
100%