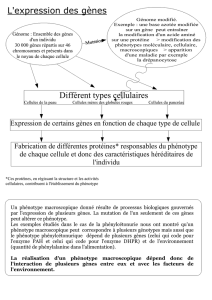
I/ La structure de l`ADN

1
Le génome est l’ensemble du matériel génétique héréditaire, il en existe deux sortes chez les
eucaryotes :
- Le génome nucléaire : les chromosomes (ADN + histones).
- Le génome mitochondrial (transmis uniquement par la mère).
I/ La structure de l'ADN :
L'acide désoxyribonucléique est composé:
- D’un support génétique 5 à 10%. (3% aboutie à la formation d’une protéine).
- D’une structure chimique.
- De régions répétées (seflish DNA) 30% du génome avec fonctions de régulation.
- De régions intergéniques, commande la transcription.
- En double brin, hélicoïdale, orienté 5’ vers 3’.
Le gène est une région d'ADN qui est susceptible d'être transcrite en ARN, ils s’expriment ou non dans
la cellule.
ADN transcription ARN traduction (région codante, ORF) protéine.
Les régions non codantes de l'ARN en amont s’appellent 5'UTR et en aval 3'UTR ne permettent pas la
traduction.
II/ La chimie de l'ADN :
Toutes les synthèses se font de 5' 3P à 3' OH.
1) Chez les procaryotes.
Chez les procaryotes la traduction et la transcription sont couplées, ARN et ADN colinéaire,
concomitantes, elles peuvent s’effectuer en même temps.
2) Chez les eucaryotes :
Il y a protection du transport de l'ARN par :
- caping, protection du 5' par G.
- polyadénylation en 3'.
- épissage (splicing) supprime les introns de l'ARNp-m ARNm.
Les introns permettent le transport ARN et le stock par fixation des protéines.
Les exons sont les parties retrouvés sur l'ARNm.
L'ARN et l'ADN sont non colinéaires.
GENETIQUE

2
La région transcrite d'ADN du gène est découplée en :
- un promoteur.
- une région transcrite.
- un signal de fin de transcription.
La transcription donne différents types d'ARN:
- ARN de structures (ribosomaux et transfert).
- ARN de régulation.
- ARNm traduit en protéine.
- ARN avec une activité enzymatique : ribozymes.
Région codante de 5' à 3' ou d’AUG au codon stop (=aucuns ARNt correspondant).
III/ La mutagénèse de l’ADN.
La réplication se fait sous l’enzyme DNA polymérase, c’est l’enzyme qui permet à partir du
triphosphate de faire l’ADN.
95% des mutations sont invisibles sur le phénotype.
On a un polymorphisme entre descendants et parents à cause des erreurs de réplication.
Les mutations moléculaires qui ont un impact phénotypique touchent :
- Les régions de régulation de la transcription (amont des promoteurs inclus).
- Les régions codantes.
- Le promoteur.
Les mutations se situent uniquement sur l’ADN.
Mutagénèse réplication de l’ADN
La réplication:
- Est semi-conservatrice, un brin parentale et un brin fille.
- Débute au niveau de séquences particulières : origine de réplication.
- La fusion de l’ADN se fait en l’origine et permet d’installer la machinerie de réplication en
fondant les liaisons hydrogènes entre les nucléotides bulle de réplication.
- Bidirectionnelle elle est continue sur un brin et est discontinue sur l’autre.
- Polymérise de 5’ vers 3’.
- Besoin d’une amorce de quelques nucléotides (contrairement à l’ARN).
- Enlève les mutations par :
Activité de relecture, l’exo nucléase 3’-5’ qui enlève le nucléotide erroné
(une erreur 108 nucléotides).
Système de réparation qui répare le nucléotide du brin transcrit ou du
brin parental.
Les agents mutagènes augmentent le taux d’erreurs de la réplication.
Dégénérescence du code génétique : plusieurs codons différents codent pour le même acide aminé,
phénomène de wobble (Gly : GGA-GGC-GGU-GGG).
Toute mutation qui remplace un codon codant pour l’acide aminé X par un codon codant pour l’acide
aminé X est qualifiée de silencieux phénotypiquement.

3
Toute mutation qui remplace un codon codant pour l’acide aminé X par un codon stop s’appelle une
mutation non-sens et le phénotype induit sera fonction de la localisation.
Toute mutation qui remplace un codon codant pour l’acide aminé X par un codon codant pour l’acide
aminé Y s’appelle une mutation faux-sens et le phénotype induit sera fonction de la nature des acides
aminés et de la position des acides aminés changés.
IV/ La génétique mendélienne.
Un gène peut correspondre à plusieurs allèles différents :
- Allèle normal (WT) : allèle le plus fréquent.
- Allèle mutant.
Un allèle muté peut induire une perte de fonction ou un gain de fonction.
Elle s’applique aux organismes diploïdes :
- une mutation récessive pas de phénotypes visible en présence de l’allèle sauvage pas
de phénotypes chez les hétérozygotes phénotypes visible chez les homozygotes.
- Une mutation dominante le phénotype s’exprime même en présence de l’allèle sauvage
chez homozygotes et hétérozygotes.
Codominance : apparition des deux phénotypes : A+B=AB.
Dominance incomplète : ni A ni B ne s’exprime mais on a un nouveau phénotype mixte C.
1) Application de la génétique mendélienne aux levures.
- Levure type a
Levure diploïde (par fusion).
- Levure type α
Le phénotype haploïde est égal au génotype.
Phénotype complexe (expérience de complémentation) :
- Générer des mutants, application agents mutagènes sur levures en division.
- Vérifier que les mutants sont récessifs, diploïde MUT + WT [WT, phénotype sauvage].
- Mélanger les mutants deux à deux (formation des diploïdes).
- Diploïdes formés les mutants affectent deux gènes différents groupe de complémentations
différents.
- Diploïdes formés les mutants affectent le même gène même groupe de complémentation.
Le nombre de groupe défini = nombre minimum de gènes impliqués dans le phénotype étudié.
2) Observations de Mendel.
La 1ère expérience de Mendel consiste a croisé deux espèces de pois l’une au grain lisse et l’autre au grain
ridé.
Il s’aperçut que ce croisement donne à 100% des pois lisse ainsi :
- Pois lisse : diploïde, homozygote, dominant, L-L, gamète L.
- Pois ridé : diploïde, homozygote, récessif, r-r, gamète r.

4
Leur croisement donne un individu diploïde hétérozygote F1 L-r, donc un pois lisse (phénotype
homogène), Dont le phénotype dépend du caractère dominant.
Une souche pure est homozygote.
Croisement de deux F1 :
- Gamète r ou L 3 génotypes :
L-r 50% pois lisse.
L-L 25% pois lisse. Phénotype dominant = ¾.
r-r 25% pois ridé.
- Mono-hybridisme :
1 gène, 2 allèles.
Pas de distorsion dans la production des gamètes.
Pas de distorsion de fécondation.
Différent phénotypiquement par un seul caractère.
Croisements de deux F0 LL-VV et rr-bb :
- Gamètes LV ou rb F1 Lr-Vb (homogène [lisse-vert] hétérozygote) après leur croisement :
L-r 50% V-b 50% (L-b=r-V) pois (lisse brun) (ridé vert)=3/16
L-L 25% V-V 25% pois lisse vert = 9/16.
r-r 25% b-b 25% pois ridé brun = 1/16.
- Di-hybridisme :
2 caractères (gènes, locus) pour chaque gène analysé les critères sont
respectés.
Les deux gènes doivent être indépendants.
(1/4) Y/Y (1/4)
LV
Lb
rV
rb
LV
LV/LV (1/16)
Lb/LV (1/16)
rV/LV (1/16)
rb/LV (1/16)
Lb
LV/Lb (1/16)
Lb/Lb (1/16)
rV/Lb (1/16)
rb/Lb (1/16)
rV
LV/rV (1/16)
Lb/rV (1/16)
rV/rV (1/16)
rb/rV (1/16)
rb
LV/rb (1/16)
Lb/rb (1/16)
rV/rb (1/16)
rb/rb (1/16)
Les phénotypes obtenus sont (allèlegène=gamète) :
- [L,V]=9/16.
- [L,b]=3/16.
- [r,V]=3/16.
- [r,b]=1/16.
Exception aux lois de Mendel le multi-allélisme, mais peut être rattaché aux expériences de Mendel en
considérant les allèles 2 par 2.
Une souche inconnue avec une souche pure récessive donne une génération au phénotype de la
souche inconnue et son génotype par la composition allélique de celle-ci test cross.

5
Exemple d’exception :
F0 mouches yeux rouges(XrXb) x mouches yeux blancs(XbY).
F1 100% yeux rouges
Toutes les XX rouges.
50% XY rouges 50% XY blancs.
XX/XY
Xr
Y
Xr
XrXr
XrY
Xb
XbXr
XbY
Les lois de Mendel ne s’appliquent pas lorsque les gènes étudiés sont liés entre eux génétiquement,
les chromosomes sexuels excepté pour la région pseudo-autosomale de Y.
3) Liaisons et carte génétique chez les diploïdes.
Liaison physique : Deux gènes sont dits liés physiquement s’ils sont sur le même chromosome.
Deux gènes physiquement liés ne sont pas forcément liés génétiquement.
Deux gènes non liés physiquement ne peuvent être liés génétiquement.
Deux gènes liés génétiquement sont séparés de maximum 50cMo.
Liaison génétique : Probabilité de n’avoir aucuns crossing over supérieure à la probabilité d’avoir un
crossing over (représente des gènes proches sur la même molécule d’ADN).
Soient 2 gènes A et B (à deux allèles) non liés physiquement, on observe la répartition des
gamètes suivante :
AB (25%), Ab (25%), aB (25%), ab (25%).
Soient deux gènes A et B (à deux allèles) liés physiquement et non génétiquement (gamètes):
AB (25%), Ab (25%), aB (25%), ab (25%).
Soient deux gènes A et B (à deux allèles) liés génétiquement (gamètes):
Majorité de gamètes parentaux AB et ab obtenus.
Minorité de gamètes recombinants Ab et aB (association des allèles produit d’une recombinaison).
La distance entre A et B ou a et b s’exprime en centimorgan : 1cMo = 1% de gamètes recombinants.
Le centimorgan est une unité de distance que pour les gènes liés génétiquement.
Exemple, cas de non liaison génétique :
F0 : AaBb x AaBb
[AB] [AB]
F1 : 9/16 [AB] + 3/16[Ab] + 3/16[aB] + 1/16[ab]
 6
6
1
/
6
100%