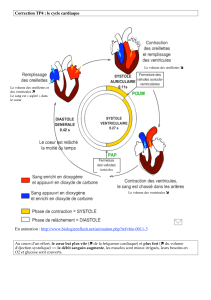
La physiologie du système circulatoire

La physiologie du système circulatoire
Le



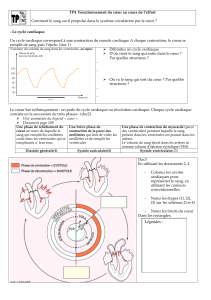
Figure 1
Le coeur assure le mouvement du sang dans la petite circulation
(VD-OG) et la grande circulation (VG-OD). Cette performance
dépend d’une organisation anatomique originale où les cavités du
coeur communiquent par différents orifices équipés de valves
antireflux, d’un système d’excitation automatique, d’une mécanique
contractile particulière, et enfin d’un apport correct en énergie et en
oxygène. ANATOMIE DU CŒUR
Le cœur est un véritable muscle creux qui est logé dans le
thorax (Figure 1).
Chez un homme au repos, le volume sanguin est en moyenne de 5 à
6 litres et le débit cardiaque d’environ 5,5 l/min. Dans l’exercice
intense le débit cardiaque peut atteindre 25 l/min. Pendant toute la
vie de l’homme, cette petite pompe pesant environ 300 grammes, «
de la taille d’un poing fermé » est responsable de la circulation.
Aucun ingénieur n’a encore su réaliser une pompe capable de durer
aussi longtemps. En 70 ans le volume éjecté par les deux ventricules
dépasse 400 millions de litres même si le sujet reste au repos.
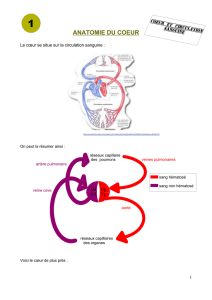
Le cœur est formé de deux moitié distinctes sans communication, le
coeur droit et le coeur gauche, séparés par une cloison médiane
étanche. Chaque moitié comprend 2 cavités : l'oreillette en haut et le
ventricule en bas (Figure 2).

Le cœur comporte quatre cavités : deux oreillettes qui, bien que
contractiles, ont surtout un rôle de réservoir et deux ventricules qui
représentent la pompe proprement dite. Oreillette et ventricule du
côté droit sont séparés de leurs homologues du côté gauche par une
paroi inter auriculaire et une paroi inter ventriculaire.
Figure 2
Ces cavités communiquent deux à deux par des orifices où le
passage du sang est rythmiquement autorisé ou interdit par des
valvules, petits clapets dont la partie mobile est reliée par des piliers
d'attache à la paroi du coeur, et que la seule pression du sang
circulant ferme de force ou laisse s'ouvrir.
HISTOLOGIE
Le coeur est constitué de trois tuniques : le péricarde, une
tunique séreuse, à l'extérieur ; le myocarde, au centre, une tunique
musculaire épaisse ; et l'endocarde, une muqueuse à l'intérieur.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
1
/
77
100%