Biochimie

Biochimie 19/11/2004 Emilie Couderc
Manon Béchu
A partir de la page.8
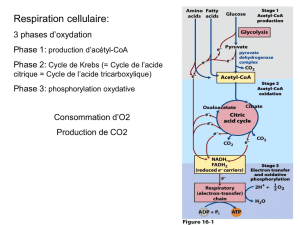
3)) CATABOLISME DES ACIDES GRAS :
1)Les acides gras, stockes dans les triglycérides, représente le plus
grand potentiel énergétique de l’organisme :
Questions :
1. En assimilant G à H, calculer le rendement :
Rendement = (129 * 7.3)/2340 = 40 %
2. Connaissant les asses molaire respectives du glucose (180) et de l’acide palmitique
(256) , comparer les valeurs énergétiques e ces deux types d’aliments :
Le glucose a une valeur énergétique de 3.8 kcal/g, tandis que l’acide palmitique a une valeur
énergétique de 9.1 kcal/g
La valeur energetique des acides gras est donc la plus importante
2) L’utilisation des acides gras comme source d’énergie varie avec les
tissus et les conditions physiologiques et pathologiques :
parmis les exemples on peut rajouter que dans le cœur il y a de nombreux LPL
3) Le transport des AG jusqu’à la mitochondrie : albumine, FABP,
activation en acyl-CoA, rôle de la carnitine :
A partie des TAG on obtient des AGL qui vont être capturés par la cellule
Ils s’intègrent dans le feuillet externe de la membrane plasmique et par flip flop, passent au
niveau du feuillet interne
Ils seront donc pris en charge par des FABP et transformés en acyl_CoA par une thiokinase
ou acétyl CoA synthétase
Ensuite en acyl-carnitine
Il y a ainsi intervention de la carnitine pour faire pénétrer les AG dans la mitochondries

Thio kinase :
Dans l’acyl-CoA il y a une liaison thio-ester et dans l’acyl-carnitine il y a une liaison de type
ester
++++ Toute réaction qui génère d l’AMP, consomme deux molécules d’ATP
GLC
malonyl_CoA
AG
L’acyl Coa est beaucoup plus volumineux que l’acyl-carn et il existe un translocateur
spécifique de l’acyl-carn = CACT, alors que l’acyl-CoA ne peut traverses la membrane
mitochondriale
La CARN s’équilibre avec le pool externe
Il y a deux iso formes de laCPT : CPT1 et CPT2 La réaction évolue donc de la gauche vers la
droite mais aussi de la droite vers la gauche
Bilan énergétique de la beta-oxydation :
Acide myristique : 14C
Il y a 7 tours de cycle de Krebs et donc 7 acétyl-CoA
Or 1 acétyl-CoA = 12 ATP
12*7= 84 ATP
De plus a chaque cycle de beta_oxydation i y a 5 ATP et comme il y a 6 tours = 30 ATP
84 + 30 = 114 -2 = 112 ATP
Remarques :
Les AG a chaînes courtes peuvent traverser la membrane mitochondriale par diffusion
Les AG a chaînes longues non et utilisent donc les peroxysomes
Donc si déficit en peroxydases, il y aura accumulation d’AG à longues chaînes
5) Cétogenèse :
Il y a u problème :
L’acetyl-CoA s accumule et ne sera pas entièrement oxyde
Quand pas suffisamment de pyruvate, on ne peut prendre en compte tout l’acétyl-CoA
PU + 10 acétyl-coa = 1 molécule de PU 1 molécule d’OA
On brûle donc une d’acetyl-CoA et il en reste donc 9 !!
La cétogenèse est donc une conséquence de l’accumulation d’acetyl-CoA qui ne peut être
consomme par le cycle de Krebs
1
/
2
100%