PHYSIOLOGIE RENALE

PCEM 2 PURPAN
PHYSIOLOGIE
RENALE
Professeur J.L. ADER

PHYSIOLOGIE RENALE PCEM2
2
SOMMAIRE
INTRODUCTION
A – RELATIONS ENTRE STRUCTURES ET FONCTIONS RENALES
B – BINEPHRECTOMIE EXPERIMENTALE ET INSUFFISANCE RENALE
C – FONCTIONS DU REIN
1 – EPURATION SELECTIVE
2 – REGULATION HOMEOSTASIQUE DU MILIEU INTERIEUR
3 – FONCTION « ENDOCRINE »
4 – PARTICIPATION A LA REGULATION DE LA PSA
II – METHODES D’ETUDE ET MECANISMES DES TRANSFERTS
NEPHRONIQUES
A – DEFINITIONS
B – METHODES D’ETUDE
1 – METHODES GLOBALES
a – METHODE DES CLEARANCES
b – MESURE DES TRANSFERTS TUBULAIRES MAXIMAUX (Tm)
c – EXCRETIONS ET REABSORPTIONS ABSOLUES ET
FRACTIONNELLES
2 – METHODES EXPERIMENTALES
a – METHODE DE DIURESE INTERROMPUE OU STOP-FLOW
b – MICROELECTRODES
C – MECANISMES
1 – TRANSFERTS PASSIFS
2 – TRANSFERTS ACTIFS
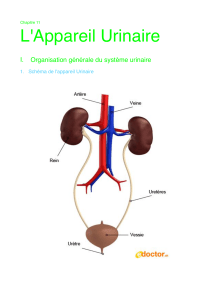
III – CIRCULATION RENALE
A – PARTICULARITES DE LA DISPOSITION VASCULAIRE
B – DEBIT SANGUIN RENAL (DSR) ET DEBIT PLASMATIQUE RENAL (DPR)
1 – MESURE DU DPR TOTAL : CLEARANCE DU PAH
2 – EVALUATION DE LA DISTRIBUTION INTRA-RENALE DU DPR ET DU
DSR
a – CLEARANCE ET EXTRACTION DU PAH
b – AUTRES TECHNIQUES

PHYSIOLOGIE RENALE PCEM2
3
c – RESULTATS
3 – CONSOMMATION D’OXYGENE
C – HEMODYNAMIQUE RENALE : PRESSIONS SANGUINES ET
RESISTANCES VASCULAIRES A L’ECOULEMENT
D – REGULATION
1 – AUTOREGULATION RENALE
2 – REGULATION EXTRINSEQUE
a – SYSTEME NEURO-ADRENERGIQUE
b – SYSTEME RENINE-ANGIOTENSINE
IV – FILTRATION GLOMERULAIRE
A – URINE PRIMITIVE (Up) : ULTRA-FILTRAT PLASMATIQUE
B – PRESSION DE FILTRATION
C – PERMEABILITE DU FILTRE GLOMERULAIRE
D – DEBIT DE FILTRATION GLOMERULAIRE (DFG)
1 – METHODES DE MESURE : CLEARANCE A L’INULINE, DE LA
CREATINE ENDOGENE ET AUTRES METHODES
2 – RESULTATS
E – REGULATION
F – ALTERATIONS DE LA FILTRATION GLOMERULAIRE
V – TRANSFERTS TUBULAIRES
A – GENERALITES
1 – BASE STRUCTURALE
2 – DEMONSTRATION : COMPARAISON FILTRATION-EXCRETION
3 – CONSEQUENCE GENERALE ET RÔLE : LA REABSORPTION
TUBULAIRE VARIABLE EST LE MECANISME HOMEOSTASIQUE
B – REABSORPTION ACTIVE
1 – GLUCOSE
2 – ACIDES AMINES
3 – ACIDE URIQUE
C – REABSORPTION PASSIVE : UREE
D – SECRETION ACTIVE : PAH
E – SECRETION PASSIVE : BASES ET ACIDES AMINES
VI – EXCRETIONS RENALES DU SODIUM ET DE L’EAU
A – BILANS DU SODIUM ET DE L’EAU

PHYSIOLOGIE RENALE PCEM2
4
B – TRANSFERTS NEPHRONIQUES
1 – SEGMENTS PROXIMAUX : GLOMERULE ET TUBULE PROXIMAL
a – SODIUM
b – EAU
2 – SEGMENTS DISTAUX : « DILUTION-CONCENTRATION »DES URINES
a – ANSE DE HENLE
– BASES
– STRUCTURALE
– ECOLOGIQUE
– PHYSIOLOGIQUE : GRADIENT D’OSMOLARITE
CORTICO-PAPILLAIRE
– CREATION ET ENTRETIEN DU GRADIENT D’OSMOLARITE
b – TUBULE CONTOURNE DISTAL ET CANAL COLLECTEUR :
UTILISATION DU GRADIENT D’OSMOLARITE
C – REGULATION DES TRANSFERTS
1 – REGULATION PROXIMALE : EQUILIBRE GLOMERULO-TUBULAIRE
2 – REGULATION DISTALE
a – SODIUM : ROLE DE L’ALDOSTERONE
b – EAU
– VARIATIONS DU GRADIENT D’OSMOLARITE
– ROLE DE L’ADH
3 – FACTEUR NATRIURETIQUE AURICULAIRE
D – RESULATS
1 – SODIUM
2 – EAU
E – EXPLORATION
VII – EXPLORATION RENALE DE POTASSIUM
A – BILAN DU POTASSIUM : BALANCES EXTERNE ET INTERNE
B – TRANSFERTS NEPHRONIQUES : FILTRATION GLOMERULAIRE,
REABSORPTION TUBULAIRE, SECRETION TUBULAIRE
C – REGULATION DE L’EXCRETION : APPORTS DE POTASSIUM, APPORTS
SODES, ETAT ACIDO-BASIQUE, ALDOSTERONE
VIII – EQUILIBRE ACIDO-BASIQUE
A – STABILITE DU Ph ET FACTEURS DE DESEQUILIBRE ACIDO-BASIQUE

PHYSIOLOGIE RENALE PCEM2
5
1 – METABOLISME OXYDATIF
2 – ABSORPTION ALIMENTAIRE
3 – SECRETIONS DIGESTIVES
B –SYSTEMES TAMPONS
1 – TAMPONS INTRA-CELLULAIRES ET OSSEUX
2 – TAMPONS EXTRA-CELLULAIRES
a – PROTEINES PLASMATIQUES ET PHOSPHATES
b – ACIDE CARBONIQUE – BICARBONATES
C – REGULATION RESPIRATOIRE
D – REGULATION RENALE
1 – MECANISME TUBULAIRE DE SECRETION D’ION H+
2 – CONSERVATION DES BICARBONATES FILTRES
a – TRANSFERTS NEPHRONIQUES
b – REGULATION
3 – REGENERATION DES BICARBONATES AVEC EXCRETION D’IONS
ACIDES
a – EXCRETION D’IONS H+ LIBRES : Ph URINAIRE
b – EXCRETION D’ACIDITE TITRABLE
c – AMMONIURIE
E – EXPLORATION
1 – pH URINAIRE
2 – DEBIT URINAIRE DES IONS H+
3 – ANIONS PLASMATIQUES INDOSES
4 – EPREUVES DYNAMIQUES DE SURCHARGE ACIDE ET ALCALINE
F – DESEQUILIBRES ACIDO-BASIQUES
1 – ACIDOSES ET ALCALOSES RESPIRATOIRES ET METABOLIQUES
2 – DIAGRAMME DE DAVENPORT
3 – COMPENSATION ET CORRECTION DES DESEQUILIBRES
IX – REGULATION RENALE DE LA PSA
A – SYSTEME RENINE-ANGIOTENSINE
B – FONCTION D’EXCRETION : RELATION PRESSION-
HYDRONATRIURESE
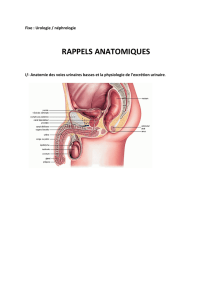
X – CONTINENCE ET MICTION
A – STRUCTURES ET INNERVATION
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
1
/
40
100%