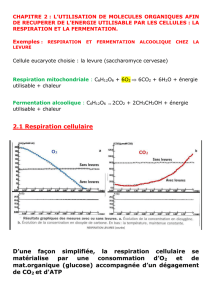
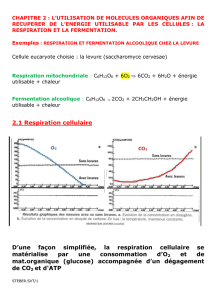
Respiration Mitochondriale - CHUPS – Jussieu

Université Pierre et Marie Curie
Respiration Mitochondriale
Objectifs au cours de
Révisions Biochimie PCEM2
Révisions Biochimie Métabolique
2004 - 2005
Pr. A. Raisonnier ([email protected])
Mise à jour : 25 novembre 2004
Relecture : Pr. A. Raisonnier

2/122 Respiration Mitochondriale - Pr A. Raisonnier 2004 - 2005

Plan du cours
2004 - 2005 Respiration Mitochondriale - Pr A. Raisonnier 3/122
Plan du cours
3Plan du cours
7Objectifs
9Partie I : Les coenzymes transporteurs d’énergie
11 Chapitre 1 : Le coenzyme ATP/ADP
12 1.1 Adénosine TriPhosphate
13 1.2 ATP → ADP
14 1.3 ATP → AMP
15 1.4 Pyrophosphatase
16 1.5 Rôles de l’ATP
17 1.6 Exemple d’ATP donneur de phosphate et d’énergie : l’hexokinase
18 1.7 Exemple d’ATP donneur de phosphate et d’énergie : la myokinase
19 1.8 Exemple d’ATP donneur d’AMP et d’énergie : l’acyl thiokinase
21 Chapitre 2 : Le coenzyme GTP/GDP
22 2.1 Guanosine Tri Phosphate
23 2.2 Rôles du GTP
24 2.3 Exemple de GTP transporteur d’énergie : la succinyl thiokinase
25 Chapitre 3 : Réactions couplées par transfert de phosphate
26 3.1 Réaction couplée
27 3.2 Créatine PhosphoKinase
28 3.3 Phosphoglycérate kinase
29 3.4 Pyruvate kinase
30 3.5 Succinyl thiokinase
31 3.6 Myokinase (adénylate kinase)
33 Partie II : Les oxydations
35 Chapitre 4 : Couples d’oxydoréduction
36 4.1 Oxydoréductions minérales
37 4.2 Potentiel standard = E’0

Plan du cours
4/122 Respiration Mitochondriale - Pr A. Raisonnier 2004 - 2005
38 4.3 Réaction couplée
39 4.4 Réaction d’oxydoréduction couplée
40 4.5 Energie d’oxydation
41 Chapitre 5 : Exemples de couples d’oxydoréduction
42 5.1 Acétaldéhyde/Acétate
43 5.2 Eau/Oxygène
44 5.3 Nicotinamide Adénine Dinucléotide
45 5.4 NADH/NAD+
46 5.5 Spectre UV du NAD+/NADH
47 5.6 Exemple de NAD transporteur d’hydrogène : l’alcool déshydrogénase
48 5.7 Exemple de NAD transporteur d’hydrogène : la malate déshydrogénase
49 5.8 Motif Rossmann
50 5.9 Flavine MonoNucléotide
51 5.10 Flavine Adénine Dinucléotide
52 5.11 Flavine réduite/Flavine
53 5.12 Flavine liée
54 5.13 Flavoprotéines
55 5.14 Coenzyme Q10 = Ubiquinone 50
56 5.15 Ubiquinol/Ubiquinone
57 5.16 Cytochrome c
58 5.17 Hème
59 5.18 Cytochromes
61 Partie III : La chaîne respiratoire mitochondriale
63 Chapitre 6 : Chaîne Respiratoire Mitochondriale = CRM
64 6.1 Complexe I
65 6.2 NADH-CoQ oxydoréductase (complexe I)
66 6.3 Complexe III
67 6.4 CoQ-cyt c oxydoréductase (complexe III)
68 6.5 Complexe IV
69 6.6 Cytochrome oxydase (complexe IV)
70 6.7 Complexe F0-F1
71 6.8 ATPase mitochondriale (complexe F0-F1)
72 6.9 Complexe II
73 6.10 Succinate déshydrogénase (complexe II)
74 6.11 ATP/ADP translocase
75 6.12 Réactions des cinq complexes

Plan du cours
2004 - 2005 Respiration Mitochondriale - Pr A. Raisonnier 5/122
77 Chapitre 7 : Bilans et schémas généraux de la CRM
78 7.1 Mitochondrie
79 7.2 Chaîne Respiratoire Mitochondriale (schéma général)
81 7.3 Membrane interne
82 7.4 Régulation par l’ADP
84 7.5 Chaîne respiratoire mitochondriale (définition)
85 7.6 Oxydation du NADH (bilan)
86 7.7 Oxydation du succinate (bilan)
87 7.8 Oxydation phosphorylante du NADH (bilan)
88 7.9 Oxydation phosphorylante du succinate (bilan)
89 Chapitre 8 : Inhibiteurs et découplants de la CRM
90 8.1 Effecteur
91 8.2 Inhibiteurs de la chaîne respiratoire mitochondriale
92 8.3 Exemple d’inhibiteur de la CRM : l’antimycine
93 8.4 Exemple d’inhibiteur de la CRM : la roténone
94 8.5 Exemple d’inhibiteur de la CRM : le malonate
95 8.6 Exemple d’inhibiteur de la CRM : le cyanure
96 8.7 Exemple d’inhibiteur de la CRM : l’oligomycine
97 8.8 Exemple d’inhibiteur de la CRM : l’atractylate
98 8.9 Exemple d’inhibiteur de la CRM : le bongkrékate
99 8.10 Découplant
100 8.11 Découplants de la chaîne respiratoire mitochondriale
101 8.12 Exemple de découplant de la CRM : le 2,4-DiNitroPhénol
102 8.13 Exemple de découplant de la CRM : l’arséniate
103 8.14 Protéines découplantes
105 Chapitre 9 : Théorie chimio-osmotique (Mitchell)
106 9.1 Gradient chimio-osmotique
107 9.2 Niveaux d’énergie de la chaîne (I)
109 9.3 Niveaux d’énergie de la chaîne (II)
111 Partie IV : Navettes mitochondriales
113 Chapitre 10 : La navette malate/aspartate
114 10.1 Malate déshydrogénase
115 10.2 Aspartate aminotransférase = ASAT
116 10.3 Navette malate-aspartate (schéma général)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
1
/
122
100%