feuille de style

La vision
Optique visuelle
Voir TD
Anatomie macro et microscopique des centres nerveux
C’est la 1ere sensibilité de l’homme
Centre périphérique : la rétine
Pour étudier cette fonction, on injecte de la proline radioactive dans l’œil. Cet acide aminé
est incorporé dans la rétine est passe dans tout le système, nous permettant de voir
l’intégralité des structures mises en jeu. L’information passe par le nerf optique, tractus
optique, corps genouillé latéral, et va jusqu’au lobe occipital.
Les neurones photorécepteurs, et les neurones de soutien sont nécessaires à la vision. Ces
neurones représentent une organisation laminaire en 8 couches.
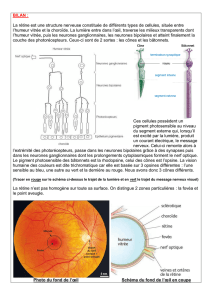
La rétine est une couche mince assez transparente qui tapisse le fond de l’œil. Cette fine
couche et composé de 8 couches qui sont en 3 parties : couche de cellule ganglionnaire,
couche de cellule interne et couche de cellule externe.
En partant du centre de l’œil à la périphérie il y a des couches de fibre optiques (terminaison
axonale), ces axones viennent des neurones placés sur la couche des cellules
ganglionnaires, puis il y a la couche des cellules bipolaires. Entre ces cellules
ganglionnaires, et les cellules bipolaires, il y a une couche plexiforme, ce sont des régions de
contacts synaptiques. Dans cette couche on va trouver des cellules de petite taille n’ayant
pas vraiment de terminaison : cellule amacrine. On trouve aussi au niveau de la couche
plexiforme externe des cellules dites horizontales.

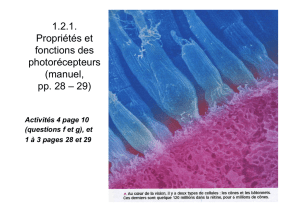
Il y a la couche des cellules photoréceptrices, car on trouve les corps des photorécepteurs,
du point de vue anatomique, il y en a deux types :
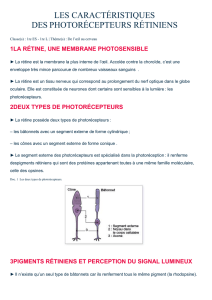
• Les cônes
• Les bâtonnets
Cônes et bâtonnets
Dans la rétine se trouvent des cellules nerveuses réceptrices spécialisées : les cônes et les
bâtonnets, qui contiennent des substances photosensibles. Ces cellules nerveuses sont nommées
ainsi en raison de leur forme.
Les bâtonnets mesurent environ 0,06 millimètres de longs et 0,25 millimètres d'épaisseur. Les cônes
sont plus courts et plus larges. Il y a environ 120 millions de bâtonnets dans un œil. Ils fonctionnent
lorsque la lumière est faible et perçoivent le noir et le blanc.
Il y a environ 7 millions de cônes dans chaque œil. Ceux-ci fonctionnent en pleine lumière. Ils
permettent de voir les couleurs. Les cônes contiennent un pigment appelé rhodopsine, qui est
décomposé et décoloré à la lumière. Ce procédé de décomposition crée un potentiel électrique qui
transforme l'énergie lumineuse en impulsion nerveuse, cette impulsion est transmise au cerveau par
l'intermédiaire du nerf optique. Ces impulsions sont interprétées par le cortex visuel pour nous
permettre de voir.
L'achromatopsie est due à des défauts des cônes. Différentes catégories de cônes sont sensibles à
différentes couleurs : les daltoniens peuvent percevoir certaines couleurs et pas d'autres. Le
daltonisme est héréditaire et est rare chez les femmes. Une alimentation correcte est essentielle pour
garder une bonne vue.
Les cônes et les bâtonnets emploient un dérivé de la vitamine A pour convertir l'énergie lumineuse en
impulsions nerveuses. La vitamine se combine à une protéine pour synthétiser un pigment
photosensible dans les bâtonnets et à la rhodopsine dans les cônes. Les personnes qui ont une
alimentation trop pauvre en vitamine A ont une mauvaise vision nocturne.
Ce lieu est appelé couche des segments externes des photorécepteurs, puis au-delà, il y a
une zone très noire appelée épithélium pigmentaire.
Les axones des cellules ganglionnaires se projettent au niveau du cerveau antérieur. Les
cellules horizontales et les cellules amacrine traitent l’information, les 3 autres types servent
à conduire l’information.
Différence régionale de la rétine : Fovéa Périphérie
Nous allons séparer 2 régions que l’on appelle Fovéa, et l’autre que l’on appelle périphérie.
La Fovéa est la partie de la rétine qui se trouve à la verticale de l’autre côté ou entre la
lumière dans l’œil. Toute la lumière qui entre dans l’œil tombe sur cette partie. Autour de
cette partie, on trouve une périphérie à 360°. La Fovéa présente 2 caractéristiques. La rétine
à cet endroit est en invagination (plus fine). La population en photorécepteur est
exclusivement formée de photorécepteurs à cône. Cela va définir la qualité de notre vision.
La lumière doit parcourir toutes les cellules avant d’arriver sur les récepteurs (cônes et
bâtonnet).
Sur la Fovéa, il n’y a que des cônes.
Nous avons environ 125 millions de photorécepteurs, nous avons un rapport bâtonnet/cône
de 20/1.

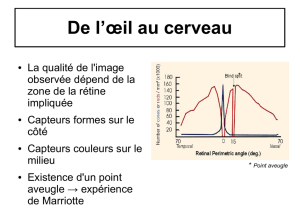
Il y a une partie de la rétine qui est dépourvue de récepteur, cette zone est située de 15°
cotés externes, on l’appelle : tache aveugle. C’est l’endroit où entrent et sortent des
vaisseaux sanguins ainsi que le nerf optique.
Les circuits rétiniens :
Nous avons 125 millions de photorécepteurs, et 1 million de cellules ganglionnaires (gros
déficit). Cela va apporter de la richesse au système, car, au niveau de la fovéa nous avons
une relation simple 1 cellule photo réceptrice pour 1 cellule bipolaire pour 1 cellule
ganglionnaire, ce rapport de 1/1/1 n’est présent que dans la fovéa. Mais vers la périphérie,
nous avons un rapport de 1000/100/1 et à l’extrémité de la périphérie nous avons un rapport
de 10000/10/1. Il y a aussi des cellules horizontales et amacrine qui sont censées être des
cellules régulatrices de l’information sensorielle.
Nerfs, Chiasma et tractus optiques :
Le nerf optique : ce sont toutes des structures axonales qui quittent l’œil pour aller se
projeter sur des structures plus profondes. Ces réseaux d’axones vont se réunir au niveau
du chiasma optique qui a une forme en H. une partie de l’information va passer dû même
coté et autre partie va passer de l’autre coté via une décussation.
Les centres intermédiaires :
Ces centres se divisent en 4 :
Noyau supra chiasmatique : voie retino-hypothalamique, cette voie informe l’hypothalamus
de la quantité de lumière présente, cela régule nos rythmes de vie.
Pretectum : voies rétino-prétectale, le prectum est situé avant le tectum, nous avons des
noyaux qui forment d’autres circuits pour réguler l’ouverture de la pupille et l’accommodation
du cristallin
Colliculus supérieur : voie rétino-tectale, la voie du colliculus, donne les informations les plus
importantes : ce sont les informations pour le mouvement des yeux et de la tête pour pouvoir
suivre un objet en mouvement. Ce sont les voies les plus connues. On a un certain nombre
de fibres qui permettent la simultanéité : synchronisme des yeux

Corps genouillés latéraux (CGP) : voie rétino-thalamique, plus de 80 % des fibres qui sortent
de l’œil. Les informations vont vers les centres supérieurs.
La représentation rétinotopique du champ visuel :
Champ de la vision binoculaire
La vision n’est pas circulaire, car la vision peut être bloquer par les pommettes, le nez,
l’arcade sourcilière… le champ visuel n’est pas quelques choses d’homogènes. On a un
champ plus étendu du côté temporal, que du côté nasal. Si on regarde avec 1 seul œil, la
partie qui est proche du nez va être regardée par la partie périphérique du côté temporal
externe.
Déficit du champ visuel
3 - Neuropathie optique ischémique antérieure :
Lier à une atteinte des petits vaisseaux nourrissant la tête du nerf optique, donnant typiquement une
hémianopsie altitudinale brutale de l’œil malade, parfois une cécité complète. Une grande
étiologie à éliminer : la maladie de Horton (plus fréquente encore que l’OACR), sinon le plus souvent
complication de l’artériosclérose.

4 - Névrites optiques
Qu’elle soit antérieure (dite papillitte, car œdème papillaire au FO) ou postérieure (dite rétro bulbaire,
car FO normal), aiguë ou chronique, uni ou bilatérale, elle donne un scotome central de l’oeil
malade (parfois les deux), et donc une baisse d’acuité Voir cours pathologie papillaire…
A titre d’exemple, la NORB unilatérale de la SEP, la NO chronique bilatérale toxique éthylo-
tabagique (cause la plus fréquente en France) ou aiguë au méthanol, la NO iatrogène à
l’éthambutol, vincristine, disulfirame…:
5- Atrophie optique compressive
Par compression orbitaire ou plus rarement par compression endocrânienne. Se souvenir (tombe en
QCM) du syndrome de Foster Kennedy qui traduit une lésion endocrânienne qui comprime le nerf
optique et entraîne une hypertension intracrânienne : on observe donc une atrophie optique du côté
de la tumeur, et un oedème papillaire par HTIC de l’autre.
2 - d Atteinte chiasmatique : demander IRM ! (Belle QR) :
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%