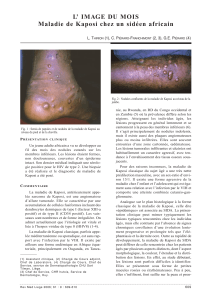
Herpesvirus associé à la maladie de Kaposi KSHV - iPubli

Il :
H à
Maladie de po

m/s • 1 1, vol. 1 1, 1!0v!e

1.
N
N
E,
1.
1
/
3
100%