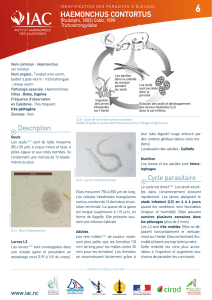
L`immunite au cours des strongyloses gastro-intestinales des

ANNEE 2002 THESE : 2002 – TOU 3 – 4077
L'IMMUNITE AU COURS DES STRONGYLOSES
GASTRO-INTESTINALES DES RUMINANTS :
Etude bibliographique
_________________
THESE
pour obtenir le grade de
DOCTEUR VETERINAIRE
DIPLOME D’ETAT
présentée et soutenue publiquement en 2002
devant l’Université Paul-Sabatier de Toulouse
par
Carine ENDERLEIN
Née, le 1er juin 1976 à FRIBOURG-EN-BRISGAU (ALLEMAGNE)
___________
Directeur de thèse : M. le Professeur Philippe DORCHIES
___________
JURY
PRESIDENT :
M. Jean-Louis FONVIEILLE
ASSESSEUR :
M. Philippe DORCHIES
M. Philippe JACQUIET
Professeur à l’Université Paul-Sabatier de TOULOUSE
Professeur à l’Ecole Nationale Vétérinaire de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE

Sommaire
1

Sommaire
2
Liste des Enseignants

Sommaire
3
Dédicaces au Jury
A Monsieur le Professeur Jean-Louis FONVIEILLE,
Professeur à l’Université Paul Sabatier de Toulouse
Praticien Hospitalier
Zoologie-Parasitologie
Qui nous a fait l’honneur d’accepter la présidence de notre jury de thèse,
Hommage respectueux.
A Monsieur le Professeur Philippe DORCHIES,
Professeur à l’Ecole Nationale Vétérinaire de Toulouse
Parasitologie et Maladies parasitaires
Qui nous a dirigé, soutenu et encouragé dans l’élaboration de notre thèse,
Qu’il trouve ici l’expression de notre vive gratitude et de notre profond respect.
A Monsieur le Docteur Philippe JACQUIET,
Maître de Conférence à l’École Nationale Vétérinaire de Toulouse
Parasitologie et Maladies parasitaires
Qui nous a fait l’honneur d’être membre de notre jury et d’examiner ce travail,
Hommage respectueux.
En témoignage de reconnaissance et de respect

Sommaire
4
Remerciements
A ma mère, à qui je dois tellement. Que ceci soit un témoignage de mon respect, de ma
reconnaissance et de tous les sentiments que je lui porte.
A ma sœur, complice et amie. Merci d’être comme tu es.
A mon père, en souvenir d’une enfance heureuse.
A Jacques et Xavier, pour leur soutien et bienvenus dans la famille.
A tous ceux qui m’ont témoigné amour et attention au cours de ces années.
A tous mes amis et mon groupe de TP, merci pour tous ces moments, bons et moins bons
passés ensembles.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
1
/
102
100%