Bulletin de conservation 2015.

BULLETIN DE CONSERVATION 2015
2016
Les parcs nous ont dévoilé...

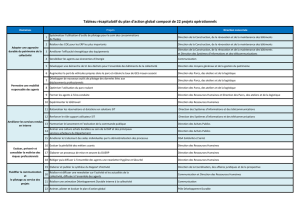
TABLE DES MATIÈRES
DE LA CONNAISSANCE À LA CONSERVATION 1
PARC NATIONAL D’ANTICOSTI 5
Mesurer l’impact du cerf de Virginie sur la flore : un projet ambitieux !
PARC NATIONAL DU FJORD-DU-SAGUENAY 8
Des carex d’exception sur les terrasses marines de Tadoussac
PARC NATIONAL DE LA GASPÉSIE 12
La fl ore exceptionnelle des hauts sommets des monts Chic-Chocs : inventaire et découvertes !
PARC NATIONAL DU MONT-SAINT-BRUNO 17
D’hier à aujourd’hui : évolution de la flore dans un parc sous pression
PARC NATIONAL DES ÎLES-DE-BOUCHERVILLE 21
Limiter l’invasion du roseau commun sur des terres en friche
PARC NATIONAL DE L’ÎLE-BONAVENTURE-ET-DU-ROCHER-PERCÉ 25
L’histoire des feux naturels sur l’Île Bonaventure
PARC NATIONAL DE MIGUASHA 27
Une vie virtuelle pour un poisson mort depuis 380 millions d’années
PARC NATIONAL DES MONTS-VALIN 31
Épidémies forestières : un plongeon dans le passé
PARC NATIONAL DES PINGUALUIT 35
Études géobotaniques au cœur de la toundra
PARC NATIONAL DE LA YAMASKA 39
Enquête sur le mercure dans la chaîne alimentaire du réservoir Choinière
PARC NATIONAL D’AIGUEBELLE 42
La protection du loup dans les parcs nationaux
PARC NATIONAL DE FRONTENAC 46
Explorer pour mieux conserver
PARC NATIONAL DU MONT-MÉGANTIC 49
Un exemple d’analyse des priorités de conservation en zone périphérique
PARCS QUÉBEC 53
Des zones périphériques au bénéfice de tous !
Photo de couverture : parc national des Grands-Jardins, Dominic Boudreault

Bulletin de conservation 2015 | 2016 1
DE LA CONNAISSANCE À LA CONSERVATION
Cette année encore, de nombreux projets de conservation ont eu lieu dans les parcs nationaux. Nous vous en présentons quelques-
uns dans cette 14e édition du Bulletin de conservation.
La réalisation de ces projets nous permet de mieux comprendre les territoires et de mieux les protéger, en nous amenant notamment
à suivre l’évolution de certaines espèces, à en découvrir de nouvelles et à lutter contre certaines plantes envahissantes.
Des études scientifiques, réalisées par des chercheurs universitaires, révèlent des pans de l’histoire naturelle du Québec ! Grâce à
elles, nous comprenons mieux le fonctionnement des écosystèmes et de leurs composantes. Tous ces projets font avancer la science
au profit de tous et contribuent à mieux préserver la biodiversité du Québec.
La conservation des parcs nationaux est tributaire de ce qui se passe en périphérie. Le dernier article de ce numéro présente les
grandes lignes d’un projet d’envergure qui s’est amorcé cet été dans le sud du Québec, en partenariat avec la Fondation de la faune du
Québec, Environnement Canada et des partenaires régionaux. Ce projet permettra de mieux connaître ces territoires et de sensibiliser
les acteurs régionaux à leur importance.
Par les diverses mesures prises depuis 1999 pour assurer la mission de conservation des parcs nationaux, la Sépaq a démontré son
engagement indéfectible à l’égard de ce motif premier de création de ces territoires.
Naturellement, nous tenons à remercier toutes les équipes des parcs qui, par leur travail et leur engagement, rendent possible ces
projets de conservation et contribuent de manière importante à l’atteinte de la mission des parcs.
Prenez quelques instants pour découvrir tout le travail qui s’accomplit pour bien connaître et protéger nos parcs nationaux québécois.
Bonne lecture !
Martin Soucy
Vice-présidence exploitation
Parcs Québec
John MacKay
Président-directeur général

4 parcsquebec.com
LES PARCS NATIONAUX DU QUÉBEC :
UN CHOIX DE SOCIÉTÉ GAGNANT !
Photo : Mathieu Dupuis
LE SAVIEZ-VOUS ?
La pratique d’activités de plein air contribue à la santé physique des gens et est un bon moyen de lutter
contre l’obésité et les maladies associées au sédentarisme.
La pratique d’activités dans la nature diminue le stress et la fatigue, et stimule les capacités mentales.
La pratique d’activités dans les parcs contribue au développement économique des régions.
Les parcs nationaux procurent des services écologiques à toute la population tels que la préservation de l’eau
potable, la purification de l’air et le maintien de populations d’espèces fauniques d’intérêt.
Les parcs nationaux contribuent au développement de l’identité des régions.
Conserver l’intégrité de ces territoires, c’est aussi assurer le maintien de ces bienfaits à long terme

Bulletin de conservation 2015 | 2016 5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
1
/
64
100%