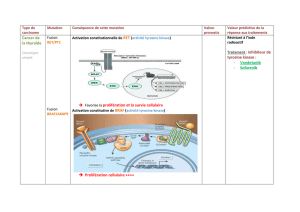
Spectre clinique des déficits en mévalonate kinase

Dossier
Spectre clinique des déficits
en mévalonate kinase
Séverine Guillaume
Service de pédiatrie générale et de rhumatologie pédiatrique, Hôpital Bicêtre,
78 rue du Général Leclerc, 94275 Le Kremlin-Bicêtre Cedex
Résumé
Les déficits en mévalonate kinase (MKD) sont des maladies rares, caractérisées
cliniquement par la survenue d’épisodes fébriles récurrents et inexpliqués chez
un enfant de moins de 3 ans, associés à divers symptômes ORL, cutanéo-
muqueux, articulaires, musculaires, digestifs et neurologiques. Ils constituent
un spectre clinique continu allant de formes modérées de syndrome hyper-IgD
aux formes gravissimes d’acidurie mévalonique. Il s’agit d’affections génétiques
autosomiques récessives, liées à des mutations du gène codant pour l’enzyme
mévalonate kinase qui intervient dans la biosynthèse du cholestérol et des
protéines isoprénylées. Aux signes cliniques s’associent en période de crise un
important syndrome inflammatoire biologique, souvent une hyper-IgD qui a
donné son nom au syndrome fébrile récurrent et une augmentation parfois
majeure de l’acide mévalonique urinaire qui permet de poser le diagnostic.
L’effondrement de l’activité mévalonate kinase dans les lymphocytes des pa-
tients permet aussi de poser le diagnostic dans les cas litigieux ; enfin, l’identi-
fication récente du gène impliqué dans ces maladies permet de rechercher les
mutations responsables de l’affection. Le traitement des MKD dépendra de la
sévérité des symptômes allant d’une corticothérapie à haute dose et courte, aux
biothérapies, exceptionnellement à la greffe de moelle osseuse.
Mots clés : mévalonate kinase, syndrome hyper-IgD, acidurie mévalonique
Les déficits en mévalonate kinase
(MKD) aussi appelés MAPS pour
mevalonate kinase associated perio-
dic syndromes, font partie du groupe
des maladies auto-inflammatoires hé-
réditaires et sont caractérisés clinique-
ment par la survenue d’épisodes fébri-
les récurrents inexpliqués, associés à
divers symptômes ORL, abdominaux,
cutanéomuqueux, articulaires et par-
fois neurologiques [1]. Les MKD ont
été essentiellement rapportés en Hol-
lande et en France, mais aussi plus
récemment en Italie, en Allemagne et
au Maghreb, avec un nombre total de
patients répertoriés de l’ordre de 210
[2, 3]. Rappelons que les MKD sont
des maladies rares, dont la prévalence
actuelle en France serait de l’ordre
d’une cinquantaine de cas. En Italie,
les MKD représenteraient 11 % des
cas de fièvres récurrentes [3]. La tra-
duction clinique des MKD revêt de
multiples aspects, qui s’inscrivent tous
dans un spectre continu de maladies,
allant de formes peu invalidantes de
syndrome hyper-IgD (HIDS) aux for-
mes létales d’acidurie mévalonique
(AM) [4, 5]. Les MKD sont dus à des
mutations dans le gène MVK qui code
m
t
p
Tirés à part : S. Guillaume
doi: 10.1684/mtp.2008.0165
mt pédiatrie, vol. 11, n° 3, mai-juin 2008
172

pour l’enzyme mévalonate kinase (MK), et sont transmis
sur un mode autosomique récessif [6, 7].
Aspects clinico-biologiques des MKD
Le syndrome hyper-IgD ou HIDS
Le HIDS débute précocement dans la vie, deux fois sur
trois avant 1 an, et dans la quasi-totalité des cas avant l’âge
de 3 ans [1, 8] ; il est constamment fait d’épisodes fébriles
récurrents et inexpliqués. Les épisodes fébriles débutent
brutalement par des frissons, des pics à 39-40 °C, et sont
souvent déclenchés par des infections mineures, les
vaccinations, des exercices physiques ou des stress émo-
tionnels. Il existe parfois une phase prodromique avec
céphalées, asthénie et irritabilité. La fièvre dure typique-
ment3à6jours et se répète à intervalles irréguliers variant
de2à6semaines [1, 3, 9].
Les poussées fébriles sont presque toujours concomi-
tantes de l’apparition d’adénopathies cervicales doulou-
reuses, d’une pharyngite, et de troubles digestifs variés
(inappétence, nausées, vomissements, douleurs abdomi-
nales, diarrhée). Parfois apparaissent une hépatospléno-
mégalie, des signes cutanés (érythème maculeux ou ma-
culopapuleux, urticaire, purpura vasculaire, érythème
noueux, aphtose buccale exceptionnellement bipolaire)
(figure 1), articulaires (arthralgies inflammatoires ou arth-
rites des grosses articulations, rarement destructrices),
musculaires (myalgies avec ou sans myolyse) ou neurolo-
giques (céphalées). Un seul cas d’orchite a été rapporté
dans la littérature au cours du HIDS [8]. Les crises fébriles
sont marquées sur le plan biologique par l’apparition
d’une hyperleucocytose à polynucléaires neutrophiles
avec monocytose et d’un syndrome inflammatoire avec
élévation majeure de la VS, de la CRP, et de la SAA
(protéine Serum Amyloid A) lorsqu’elle est dosée.
Ces paramètres inflammatoires ainsi que le tableau
clinique se normalisent habituellement entre les crises
[1, 3, 9]. En revanche, des taux parfois élevés d’IgD
persistent chez certains patients, même en dehors des
crises (0-5 300 UI/ml). Ceci a valu son nom à ce syndrome
fébrile récurrent, identifié en 1984 par le groupe de Van
der Meer, en Hollande : le HIDS, auparavant appelé ″fiè-
vre hollandaise″(dutch fever) [10]. En fait, le HIDS avait
fait l’objet un peu plus tôt d’une description dans une
publication française par Prieur et Griscelli, qui y avaient
reconnu une entité clinique à part, distincte de la maladie
de Still [11]. Malheureusement, la présence d’une hyper-
IgD n’est pas constante, ayant été retrouvée normale chez
des patients gravement atteints avec HIDS génétiquement
confirmé ; de plus, le tableau clinique complet de HIDS
peut précéder l’apparition de l’hyper-IgD de plusieurs
années [1, 7, 8, 12-14]. L’hyper-IgD n’est pas non plus
spécifique puisqu’elle est observée dans de nombreuses
autres pathologies inflammatoires chroniques [12]. La
physiopathogénie et le rôle de l’hyper-IgD ne sont actuel-
lement pas encore élucidés ; elle est donc pour le moment
considérée comme un épiphénomène, et non comme un
marqueur biologique du HIDS. Pour des raisons tout aussi
obscures, des taux élevés d’IgA1 sont observés chez plus
de 90 % des patients atteints de MKD [8]. Enfin, des études
sur les cytokines circulantes ont montré un excès de
cytokines pro-inflammatoires ex vivo (TNFa, IL-6, IL-18,
+/– IL-1b) chez les malades par rapport aux sujets sains ;
après stimulation de leurs cellules sanguines mononu-
cléées in vitro par du lipopolysaccharide (LPS), il était noté
une élévation des taux de TNFa, à un moindre degré d’IL-6
et une augmentation spectaculaire du niveau d’IL-1bdans
les cellules des patients par rapport aux sujets contrôles
[15, 16]. L’analyse du liquide céphalorachidien a été
effectuée chez certains patients et a toujours été normale.
Lorsque les arguments cliniques sont suffisamment
évocateurs de HIDS, le diagnostic peut être porté sur la
présence d’une hypermévalonaturie (excès d’excrétion
urinaire d’acide mévalonique) au cours d’un épisode fé-
brile ; c’est en effet pendant les poussées fébriles que le
déficit en MK se creuse, faisant apparaître l’acide mévalo-
nique dans les urines (taux de mévalonate urinaire variant
de 0,004 à 0,028 mol/mol de créatinine pour une norme
inférieure à 0,001) [17]. Lorsque le cas est litigieux, on
peut demander une évaluation de l’activité MK dans les
lymphocytes du malade au mieux en période fébrile.
L’activité MK dans les lymphocytes des patients avec
HIDS est en général de l’ordre de1à7%,uneactivité MK
supérieure à 30 % de la norme étant habituellement suffi-
sante pour produire la quantité de composés situés en aval
Figure 1.Lésions cutanées au cours d’épisodes fébriles de syndrome
hyper-IgD (HIDS). Photo : Frenkel, Journée du CRI 2005.
mt pédiatrie, vol. 11, n° 3, mai-juin 2008 173

de la MK nécessaires à une vie normale [18] ; les sujets
hétérozygotes ont une activité MK de l’ordre de 60 % par
rapport au témoin du jour. Enfin, l’étude du gène MVK en
biologie moléculaire, rendue possible grâce au travail de 2
équipes scientifiques hollandaise et franco-hollandaise en
1999 [6, 7], permet d’identifier les mutations responsables
du MKD. Ces mutations sont principalement faites de
mutations faux-sens (remplacement ponctuel d’un acide
aminé par un autre dans 84 % des cas) distribuées tout le
long du gène qui est situé sur le bras court du chromosome
12 (figure 2, [19]). Plus de 70 mutations de MVK ont été
identifiées, et sont répertoriées dans le site web dédié aux
syndromes auto-inflammatoires créé par Touitou et al.
[19]. Les mutations qui déstabilisent le plus la structure
tertiaire de la MK sont celles qui se traduisent par la forme
clinique la plus grave (bonne corrélation phénotype/
génotype) [20]. La plupart des malades souffrant de HIDS
sont hétérozygotes composites (doubles hétérozygotes),
c’est-à-dire qu’ils présentent une mutation différente sur
chaque allèle, et dans plus de 80 % des cas, les patients
atteints de HIDS portent une mutation V377I sur l’un des
allèles. Cette mutation a été qualifiée de « modérée »
puisqu’elle a été identifiée chez des malades souffrant
presque toujours de HIDS [4, 18]. Notons enfin que la
fréquence des individus porteurs de cette mutation en
Hollande est de 1/153 individus [21].
Il existe donc tout un continuum de formes cliniques
du HIDS, variant de l’individu asymptomatique au patient
sévèrement handicapé avec de multiples récurrences.
L’évolution de la majorité des HIDS se fait vers une
atténuation progressive des symptômes, et, dans ce cas,
est de bon pronostic ; néanmoins, il a été décrit de rares
cas de HIDS se compliquant d’amylose systémique AA,
d’insuffisance rénale terminale, de colite, de syndrome
d’activation macrophagique ou d’infections graves [3, 4,
22, 23].
Il faut se souvenir que le HIDS est un diagnostic
d’élimination d’une fièvre récidivante et qu’il est impératif
d’éliminer les causes tumorales et infectieuses de fièvre
récurrente, largement plus fréquentes, avant d’envisager
un diagnostic de HIDS.
L’acidurie mévalonique ou AM
Beaucoup plus rare que le HIDS, l’AM concernerait
une trentaine d’enfants dans le monde [2] ; elle représente
la forme grave du MKD [24, 25]. Les symptômes apparais-
sent également tôt, en général dans la première année de
vie, et se caractérisent non seulement par des poussées
fébriles irrégulières et inconstantes s’accompagnant de
malaise, d’adénopathies, d’hépatosplénomégalie, de trou-
bles digestifs parfois majeurs, d’atteintes articulaires, et de
rashs avec œdème puis atrophie du tissu sous-cutané,
mais aussi par un retard de croissance, une dysmorphie
faciale (figure 3) et des troubles neurosensoriels et muscu-
laires plus ou moins prononcés (retard psychomoteur avec
hypotonie qui reste stable dans le temps, épilepsie, ataxie
cérébelleuse progressive, hypoacousie, baisse d’acuité vi-
suelle voire cécité par cataracte, uvéite, atrophie du nerf
optique ou rétinite pigmentaire). Ces signes accompagna-
teurs des poussées fébriles régressent rarement complète-
ment entre les périodes critiques. Les formes graves d’AM
conduisent au décès de l’enfant avant l’âge de 2 ans, en
l’absence de prise en charge thérapeutique adéquate [26,
27]. En revanche, dans certaines formes atténuées d’AM,
les patients ne présentent pas de troubles psychomoteurs.
Chromosome 12q24
Gène > 18 kb 43 kDa
11 exons
ARNm 1,9 kb Protéine 397 AA K13X
H20P
H44fs
72-73insT
A141X
L234P
T243I
L264F
L265P
I268T
S272F
N301T
V310M
A334T
421-422insG
Acidurie
mévalonique #
PTS 2 ATP Binding Site
193
Del exon 2 Del exon 5Del exon 3
2013
ins ins
##
#
#
############
HIDS
#
204
H380R
R215Q
G202R
L39P
H20N
H20Q
I268T#
T209A
S272F#
R277C/H
W62X
V8L L35S
W188X
G211A
G211E
V250I
L265R S329R G376V
H20P#G309S
V377I#
C367S
N205D
P288L V293M
G335G G336S S378P
MUTATIONS du Gène MVK - Protéine MK
V203A
P167L
S135L
A147T
Y149X
I119M
V132I
G171R
A148T
A141R
T356M
T237S
L246P
T322S
G326R
Figure 2.Mutations de la mévalonate kinase (MK) dans le syndrome
hyper-IgD (HIDS) et l’acidurie mévalonique (AM).
Figure 3.Dysmorphie faciale au cours de l’acidurie mévalonique.
Photo : Frenkel, Journée du CRI 2005.
Spectre clinique des déficits en mévalonate kinase
mt pédiatrie, vol. 11, n° 3, mai-juin 2008
174

Sur le plan biologique également, il est rare que les
paramètres inflammatoires couramment retrouvés se nor-
malisent entre deux crises ; de plus, la mévalonaturie est
souvent élevée (1-56 mol/mol de créatinine pour une
norme inférieure à 0,001), et persiste entre les crises
fébriles. Lorsque l’activité MK est testée, elle est effondrée
et quasiment toujours inférieure à 1 % par rapport au
témoin du jour (en général < 0,3 %). Enfin, les IgD sont le
plus souvent très élevées chez les patients AM [2, 26].
Sur le plan génétique, il semble que certaines muta-
tions faux-sens ne soient observées que dans les AM, leur
accordant « un poids » ou une gravité particulière, par
opposition aux mutations « modérées » comme la V377I
[4, 17, 20].
L’évolution est fatale dans les formes graves d’AM, et
compliquée essentiellement de dégénérescence cérébel-
leuse, de troubles du développement psychomoteur et
staturo-pondéral, d’amylose systémique et d’infections
graves chez les patients atteints de formes moins sévères
d’AM [26-28].
Aspects physiopathogéniques
des MKD
Les mutations de MVK sont répertoriées dans le site
web dédié aux fièvres récurrentes auto-inflammatoires
Infevers
1
[19], accessible gratuitement et régulièrement
mis à jour (figure 2).
La MK est une enzyme située sur la voie de synthèse du
cholestérol et des isoprènes non-stérols, juste en aval de
l’HMG-CoA réductase. Elle est ubiquitaire et exprimée dans
les peroxysomes et dans le cytosol des cellules [29]
.LaMK
a pour fonction de phosphoryler, grâce à la présence
d’ATP, l’acide mévalonique en 5-phosphomévalonate, qui
devient à son tour le substrat d’une autre enzyme de cette
voie de biosynthèse. La structure tertiaire de cette protéine
est illustrée en figure 4A ; cette représentation est tirée de
la cristallisation de la MK de rat en présence d’une molé-
cule d’ATP et de magnésium (Mg), MK de rat qui présente
82 % d’homologie avec la MK humaine [30].
Les études
de cristallographie dévoilent les repliements en trois dimen-
sions des protéines, et permettent non seulement de situer
les acides aminés dans la structure tridimensionnelle, mais
aussi de repérer les interactions moléculaires des atomes
constituant la protéine d’intérêt. À l’état naturel, la MK
existe sous forme d’homodimère, c’est-à-dire de 2 molé-
cules de MK (figure 4B)
[30]. Elle reçoit son substrat, l’acide
mévalonique, et l’ATP nécessaire à la réaction de phos-
phorylation, dans sa poche catalytique, comme illustré
par les figure 4C et D[30]. Ainsi, on découvre dans la
figure 4A, que la valine en position 377 qui est la cible de
la mutation Valine377Isoleucine (V377I), est située en
dehors du site catalytique de l’enzyme, expliquant la
raison pour laquelle cette mutation a un impact modéré
sur le phénotype du MKD [30]. La figure 5 illustre la voie
de biosynthèse des isoprènes, et montre que le déficit en
MK induit d’une part une accumulation d’acide mévalo-
nique en amont, d’autre part un déficit en composés
d’aval [9]. Il a récemment été montré que la responsabilité
de la pathologie n’incombait pas à l’accumulation d’acide
mévalonique, mais plutôt au déficit en produits d’aval, les
isoprènes non-stérols. Les isoprènes sont des composés
essentiels, impliqués dans de nombreuses fonctions cellu-
laires, comme le transport des électrons dans la chaîne
respiratoire, la traduction des ARNm en protéines, la
glycosylation des protéines et les processus de différencia-
tion, prolifération et motilité cellulaires. Ces processus
reposent sur le fonctionnement harmonieux d’une cas-
cade enzymatique dans laquelle interviennent des GTPa-
ses cytosoliques, qui sont des petites protéines G. D’après
les travaux du groupe de Waterham en Hollande, la patho-
génie du MKD serait liée à l’absence de géranyl-
géranylation de ces petites protéines G, en particulier
celles de la famille Ras/Rho/Rac [31]. La géranyl-
géranylation consiste en l’addition d’un groupe lipidique
hydrophobique géranyl-géranyl sur une protéine cyto-
plasmique. Cette modification induit l’arrimage des pro-
téines cytosoliques à la membrane plasmique bi-lipidique
où se situent les substrats de ces enzymes, permettant le
démarrage d’une cascade de réactions enzymatiques dans
un compartiment cellulaire bien précis.
Par ailleurs, comme il l’a déjà été mentionné, il existe
dans les cellules des malades atteints de MKD, une pro-
pension à produire de l’IL-1ben excès après stimulation in
vitro [16]. Pour l’instant, le lien exact entre le défaut de
géranyl-géranylation et la production excessive d’IL-1b
dans certaines circonstances, n’est pas encore élucidé.
Des données préliminaires semblent incriminer une acti-
vation inadéquate de la caspase-1, aussi appelée ICE
(IL-1bconverting enzyme), dans la genèse de l’hyperpro-
duction d’IL-1b[32].
Enfin, il a été montré que la MK était présente dans tous
les tissus mais en quantités variables, et qu’elle était natu-
rellement thermosensible, c’est-à-dire que sa stabilité et
par conséquent son activité diminuaient chez des patients,
déjà déficitaires, lors de l’élévation de la température du
milieu ambiant. Ceci explique qu’à l’occasion d’une aug-
mentation de température, l’activité MK s’abaisse en deçà
d’un seuil critique et produise une cascade de signes
cliniques en rapport avec une vague pro-inflammatoire
[17].
La biosynthèse des isoprènes est finement régulée pour
permettre une production constante de dérivés isoprény-
lés sans accumulation de produits intermédiaires toxi-
ques, ni déficit en composés terminaux. Ainsi, le choles-
térol agit comme « senseur » de cette voie de synthèse et
exerce un rétrocontrôle inverse sur la production de MK
1
http://fmf.igh.cnrs.fr/infevers.
mt pédiatrie, vol. 11, n° 3, mai-juin 2008 175

A B
C D
H20 A334
T243
R241
S146
E193
G142
ATP
Mg
D204
K13
MEV
Figure 4.Structure de la mévalonate kinase (MK). A) Structure en 3D (hélices
␣
et feuillets b) de la MK en interaction avec une molécule
d’ATP-Mg. B) Structure de l’homodimère de MK. C) Structure présumée de la MK interagissant avec l’acide mévalonique (MEV) et l’ATP-Mg.
D) Idem C en modélisation moléculaire avec zoom. D’après Fu, J Biol Chem 2002.
Spectre clinique des déficits en mévalonate kinase
mt pédiatrie, vol. 11, n° 3, mai-juin 2008
176
 6
7
8
6
7
8
1
/
8
100%