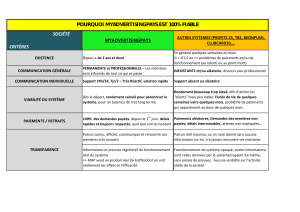
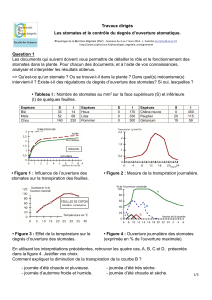
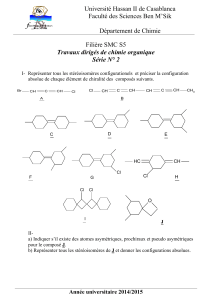
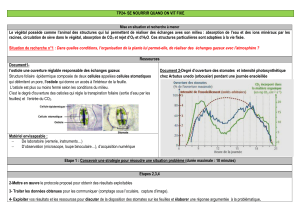
stomates_texte_anima..

Les feuilles des Plantes Supérieures disposent au niveau de leur épiderme de structures très
élaborées appelées stomates, spécialisées dans les échanges gazeux avec l’environnement et
qui leur permettent de faire face aux différentes contraintes abiotiques.
L’exemple choisi, la mise en place des stomates, résume l’ensemble des étapes et des
processus complexes qui régissent la construction de la plante. Cette mise en place procède
d’un contrôle rigoureux des processus de division cellulaire et de différenciation. Elle aboutit
à l’installation de deux cellules adjacentes, les cellules de garde du stomate. Une cellule
initiale du protoderme, par des divisions cellulaires, parfois asymétriques, établit un lignage
cellulaire dont certains éléments vont subir une différenciation.
DESCRIPTION DES DIFFERENTES ETAPES ET DE LEUR CONTROLE
plusieurs étapes, chacune sous le contrôle d’un ou de plusieurs facteurs de transcription
(Speechless, Mute, Fama et Myb88), ainsi que d’une voie de signalisation de type module
MAP kinase (Yoda, MKK4 et 5, MPK3 et 6) ont été identifiées ?
1-2 La première étape, encore mal caractérisée, produit une cellule mère du méristémoïde à
partir d’une cellule du protoderme.
2-3 La cellule mère du méristémoïde entreprend une division, asymétrique, qui génère d’une
part une large cellule épidermique et d’autre part une petite cellule, le méristémoïde.
La division asymétrique est l’un des mécanismes utilisé pour générer différents devenirs cellulaires parmi les
cellules sœurs, d’où la notion de lignage cellulaire.
Cette première étape est sous le contrôle positif de la protéine SPEECHLESS, un facteur de
transcription de type bHLH, et sous le contrôle négatif d’un module de signalisation de type
MAP kinase (comprenant YODA, MKK4-MKK5, MPK3-MPK6).
3-4 La transition suivante du méristémoïde vers la cellule mère des cellules de garde, est sous
le contrôle du facteur de transcription MUTE de type bHLH.
4-5 Enfin, la dernière étape requiert une division symétrique de la cellule mère des cellules de
garde, pour donner les deux cellules de garde. Elle met en œuvre les protéines FAMA et
MYB88. La protéine FAMA permet de déterminer l’identité « cellule de garde » et empêche
toute division cellulaire ultérieure.
Les différents gènes identifiés ont été mutés ou sur-exprimés, de façon à mieux comprendre
leur implication dans les mécanismes de division cellulaire et de différenciation.
6a-9a L’observation du mutant speechless (perte de fonction), montre que les divisions
asymétriques ne se produisent plus. l’épiderme apparaît comme un champ de cellules en
puzzle imbriquées. Le même phénotype peut être observé lorsque le module MAP kinase est
activé de façon constitutive (gain de fonction).
6b-10 La mutation du gène yoda, ou les doubles mutants mkk4-mkk5 et mpk3-mpk6 entrainent
un développement de l’épiderme qui n’est plus constitué que de cellules de garde des
stomates appariées. Le même phénotype épidermique est également observé lors de la
surexpression du gène MUTE.

9b Lorsque le gène SPEECHLESS est surexprimé, les cellules épidermiques en puzzle se
divisent de façon excessive, en exprimant un marqueur du lignage stomatique qui caractérise
habituellement les cellules mères du méristémoïde.
7 Dans le mutant mute, de nombreuses divisions asymétriques produisent une structure en
rosette, bloquée à un stade de type cellue méristémoïde
8-11 Dans le mutant fama, les cellules mères des cellules de garde se divisent plusieurs fois de
façon symétrique, sans jamais former de cellules de garde. Il en résulte une structure ovale
constituée de fines cellules alignées. Il n’existe pas de différenciation terminale. Un
phénotype du même type peut être observé chez le double mutant four lips-myb88. La
surexpression du gène FAMA produit uniquement des cellules de garde, mais celles-ci ne sont
pas appariées, donnant une structure en écailles de poisson.
Ces données permettent de dégager quelques conclusions générales concernant le mode
d’action des facteurs de transcription qui pparaissent être des acteurs positifs et le rôle de la
voie de signalisation MAP kinase qui pourrait exercer un contrôle négatif. En effet, la perte de
fonction du module MAP kinase conduit au même phénotype que la surexpression de MUTE ,
avec une accumulation de cellules de garde. Réciproquement, l’expression constitutive du
module MAP kinase (gain de fonction) et la perte de fonction du mutant speechless
produisent un épiderme constitué uniquement de cellules en puzzle.
Malgré cette accumulation de connaissances, plusieurs points demeurent à éclaircir :
- existe-t-il d’autres éléments qui interviendraient dans ce schéma ?
- quel est le lien entre les facteurs de transcription décrits et le module MAP kinase ? en sont-
ils les cibles directes ?
- quel mécanisme permet de déterminer quelle cellule sera à l’origine d’une cellule mère du
méristémoïde ? Ce mécanisme fait-il intervenir un effet de position, puisque chaque stomate
est associé à une chambre sous-stomatique ?
- comment ce mécanisme fin de régulation est-il contrôlé par l’environnement, qui est capable
de modifier considérablement le nombre de stomates par unité de surface foliaire ? Puisque
MKK4 et MKK5 jouent un rôle dans le la réponse à des contraintes environnementales, le
module MAP kinase pourrait être un acteur de ce mécanisme.
- le mécanisme qui prévaut pour les divisions asymétriques de la cellule mère du
méristémoïde, voire du méristémoïde lui-même, contrôlent-ils les autres divisions
asymétriques observées durant le développement de la plante ?
1
/
2
100%