Classification du végétal

Organisation du vivant Végétal
I] Les « mousses »
1) Caractéristiques retenues pour la classification historique
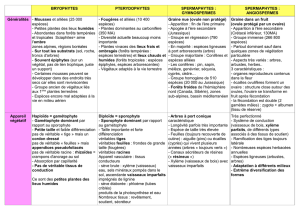
Thallophytes (plantes à thalles) Cormophytes (plantes à cormus) ou archégoniates
Cellule mère donne des spores ou gamètes fertiles « -cystes » Cellule mère donne des cellules fertiles et stériles « -anges »
Prendre schémas des spores
Les mousses sont de petite taille car elles ne possèdent pas de tissus de soutient (généralement les vaisseaux conducteurs de la
sève), donc ne peuvent pas grandir. Ce sont les plus anciens Chlorophytes terrestres (470 MA pour les plus vieux spores
trouvés, qui correspond à la conquête des espaces terrestres).
Les Bryophytes sont composés de :
•Bryopsida : mousses
•Hepatopsida : hépatiques
•Anthéceropsida : anthécérotées
2) Classification phylogénétique
Le terme mousse devient obsolète, et les Bryophytes deviennent un phylum parmi 3 phylums qui regroupent les mousses :
•Bryophytes
•Anthocérophytes
•Marchantiophytes
Embryophytes : partie de la lignée verte qui comprend les « algues vertes » (Chlorophycées)
Embryophytes
Marchantiophytes
Stomatophytes
Anthérophytes
Bryophytes
Polysporangiophytes

3) Caractéristiques « transversales »
a) Appareil végétatif
•Lames posées sur le sol
•Tiges feuillées
b) Appareil racinaire
Pas de racines, mais des rhizoïdes (soit fil de cellules, ou une seule cellule). Il n'y a pas de tissus différenciés dans un rhizoïde.
c) Différenciation cellulaire
α) Les stomates
Cellules particulières superficielle qui ont la capacité d'ouvrir ou fermer un orifice : l'ostiole.
Rôle primordial dans les échanges gazeux.
β) Les tissus conducteurs
On trouve des tissus conducteurs seulement chez les Bryophytes, mais à un stade archaïque. Les parois de ces tissus ne sont
pas lignifiées. Deux types de cellules existent : hydroïdes (conduit l'eau) et leptoïdes (conduit les nutriments). Pour les
Anthocérophytes et Marchantiophytes, il n'y a pas de tissus conducteurs.
γ) Cytologie
Anthocérophytes : 1 plaste par cellule avec pyrénoïdes (rôle dans l'anabolisme photosynthétique, notamment dans l'élongation
des sucres)
Chez les deux autres, il y a plusieurs plastes sans pyrénoïdes.
δ) Cycle de reproduction
Ce sont des cycles digénétiques haplo-diploïdes avec alternance de phase. Ce cycle correspond
à tous les végétaux de la lignée verte que l'on verra. Le gamétophyte ne ressemble pas au
sporophyte, on parle de générations hétéromorphe. Les « mousses » sont dites à gamétophytes
dominants (les mousses telles qu'on les connait sont donc haploïdes).
Ce sont des reproduction Oogames (avec oosphère et anthérozoïde nageur), qui implique donc la présence d'humidité pour
permettre aux anthérozoïdes de rejoindre l'oosphère. Le gamétophyte produit des gamétanges :
Anthéridies Archégone
Cellules stériles entourent les fertiles
Des cellules stériles entoure une oosphère
Cycles des Bryophytes
–Cycle de Bryum (Bryales)
1. Le sporophytes issus de la fécondation est hétéromorphe par
rapport aux gamétophytes.
2. Il émet des spores identiques (isosporie) qui font donner des
protonéma (filaments) qui formeront les gamétophytes
3. Les gamétophytes émettent anthérozoïdes
4. Les anthérozoïdes fécondent les archégones
5. L'œuf se divise par mitose
6. Développement du sporophyte qui est un adelphoparasite du
gamétophyte.
Tissu Stérile
Tissu fertile :
Sporgangifère
Suçoir :
Haustorium

Sphaignes (Sphagnales) Andréa (Andréales)
Cycle des Marchantiophytes (ci-contre : hépatiques)
C'est une oogamie classique. Après fécondation, l'œuf se divise par
mitose, et le sporophytes (capsules sporangifères) croit jusqu'à
éclater (déhiscence : ouverture spontanée d'organes chez les
végétaux), laissant s'échapper les spores, qui reformeront les
gamétophytes.
Jungermannia
(Jungermanniales)
L'appareil végétatif est prostré (sur le sol) et feuillé, l'archégone est dans le sol, dans une
sorte de poche : le marsupium. Une fois fécondé, l'œuf forme le sporophyte avec déhiscence
classique.
Cycle des Anthocérophytes
Reconnaissables par leurs unique plaste avec pyrénoïdes.
Les gamétophytes sont dans des cryptes à l'intérieur de la
plante. Une fois fécondés, le sporophytes sort (sous forme
de tube) et la déhiscence se fait par fente de ce tube.
3) Conclusion
Toutes les mousses ont un cycle digénétique à gamétophytes dominant et pratiquent l'oogamie. Le sporophyte est toujours
un adelphoparasite du gamétophyte femelle, et les spores générés sont tous identiques (isosporie).

II] Les « fougères », les ptéridophytes
Dans le classification historique seulement. Ce groupe n'existe plus dans la classification phylogénétique.
1) Caractéristique de la classification historique
Elles n'ont pas de fleurs et sont donc des cryptogames. Ce sont des plantes à
cormus : des cormophytes. Elles ont des feuilles, avec un système végétatif
vasculaire qui joue également le rôle de soutient. C'est ce système vasculaire à
parois lignifiées qui est une innovation dans le végétal, et sont arborescente grâce
à cet appareil (ce sont donc des Trachéophytes). Elles tirent donc leur nom de
cryptogames vasculaires de ces caractéristiques.
2) Classification phylogénétique
Ce sont des polysponrangiophytes
3) Caractéristiques transversales des fougères
a) Feuilles
Absence de feuille chez les Rhyniophytes (microphyles)
Feuilles archaïques chez Lycophytes
Feuilles normales chez les autres fougères (mégaphyles)
La différence entre microphyles et mégaphyles se situe au niveau des vaisseaux conducteurs de la sève :
Chez les mégaphyles, les stèles sont discontinues au niveau du départ des feuilles.
b) Racines
Pas de racine chez les Rhyniophytes. Toutes le fougères que l'on voit sont des
sporophytes. Tous les sporophytes des autres groupes possèdent des racines.
Polysporangiophytes
Lycophytes
Euphylophytes
Moniliformes
Spermatophytes
Filicophytes
Sphénophytes
Polysporangiophytes
Paratrachéophytes
Ou Rhyniophytes
Trachéophytes

c) Croissance et ramification de la tige
Les tiges peuvent être prostrées et souterraines, on parle alors de rhizomes. De cette tiges, plusieurs feuilles
sortent du sol (ou frondes)Les tiges peuvent être ramifiée (dichotome), ou verticillés (notamment pour la prêle
à gauche).
d) L'appareil vasculaire
On appelle stèle l'ensemble des cellules qui constitue l'appareil vasculaire. Les cellules de cet
appareil sont lignifiées (donc rigides et hydrophobes), et ont perdu leur noyau.
Le xylème transporte la sève brute chargée en eau et minéraux dans la plante (vers le haut). Ces
cellules sont très lignifiées, on parle de bois.
Le phloème transporte la sève élaborée, et le flux est descendant.
Moins lignifiées que celles du xylèmes, on parle de liber et ces
cellules sont dites criblées.
On trouve plusieurs types de stèles :
Les protostèles Les siphonostèles Les actinostèles
Presque toutes
e) La reproduction
α) Les sporanges
Les fougères ont besoin d'eau pour se reproduire et font la polysporangie. Le cycles sont digénétique à sporophytes dominant.
Chez les eusporangiés, les sporanges se développent à partir d'un groupe de cellule épidermique indifférencié et le tapis
nourricier se fait à partir de cellules sporogènes devenues stériles (tapetum), et le nombre de spores produits par de tels
sporanges est extrêmement élevé (de 15000 à 500 millions)
Chez les leptosporangés, le sporange se développe à partir d'une seule cellule épidermique. Le tapis n'est formé que par le tissu
sporogène, et le nombre de spore est très faible (minimum 4).
β) Les spores
Les fougères font généralement de l'isosporie : tous les spores sont identiques. Certaines fougères font de l'hétérosporie,
différenciés notamment par la taille (microspore et mégaspore). Les spores deviennent élaborés notamment grâce à leur
protection : membrane puis intine puis exine, puis périspore.
γ) Cycles de reproduction
Le gamétophyte est initialement assez développé pour contenir des vaisseaux conducteurs. Dans un premier temps, ce
gamétophyte était dominant. Au cours de l'évolution, le sporophyte est devenu indépendant (notamment grâce à sa résistance
due à la diploïdie, et à la perte de la dépendance parasitaire au gamétophyte) pour devenir au final plus grand et dominant.
En parallèle, les gamétophytes ont régressé par leur taille et ont perdu les vaisseaux conducteur (on parle d'involution) pour
devenir thalle (appareil végétatif très simple) ou minuscule tubercules au ras du sol (on parle de prothalle). Généralement
l'isosporie est associé à l'isoprothallie (thalles identiques et monoïques). On remarque que monoécie des pieds ne rime pas
forcément avec monoécie des fleurs (fleurs peuvent être dioïques sur des pieds monoïques).
En cas d'hétérosporie, on a des plantes dioïques, avec généralement des petits spores donnant des petits thalles mâle, et des
gros spores donnant des gros thalles femelle.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%