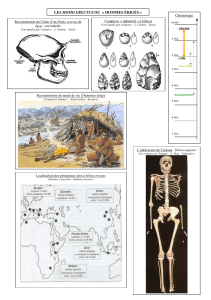
Chapitre 4 - Sciences de la Vie et de la Terre, Biologie Géologie

Chapitre 4 : Un regard sur l’évolution de l’Homme
L’origine de l’humanité ? On sait depuis longtemps par des évidences paléontologique, archéologique et biologique,
que l’homme anatomiquement moderne, Homo Sapiens, est le résultat de l’évolution d’une des branches de l’arbre
de la vie, et qu’il a ainsi des liens de parenté avec les grands singes. De plus, le rameau humain porte plusieurs
espèces aujourd’hui disparues, telles que l’homme de Néandertal ou les australopithèques. Depuis une quinzaine
d’années, la paléontologie humaine s’est enrichie de découvertes nombreuses, associée à la paléogénétique, qui ont
fortement modifié la vision de l’évolution de l’homme.
Homo sapiens peut être regardé, sur le plan évolutif, comme toute autre espèce. Il a une histoire évolutive
et est en perpétuelle évolution. Cette histoire fait partie de celle, plus générale, des primates.
Problème posé : Comment Homo sapiens, en tant que primate, peut-il être regardé scientifiquement sur le
plan évolutif ?
I. Quelles sont les relations de parenté entre l’Homme et d’autres primates et comment expliquer les
grandes différences phénotypiques entre l’Homme et ces primates ? TP n°6
1. L’Homme primate Hominidé
Le groupe des Primates contient toutes les espèces de Mammifères qui se distinguent par différents caractères
dérivés : le pouce de la main plus court et opposable aux autres doigts rendant la main préhensile, main et pied
possèdent des ongles (et non des griffes), yeux à orbites larges orientés vers l’avant permettant une excellente
vision, cerveau relativement important par rapport au corps avec un cortex bien développé.
L’état dérivé d’un caractère (par opposition à l’état ancestral) est celui apparu le plus récemment au cours des
temps géologiques. Les êtres vivants qui partagent un même caractère dérivé ont un ancêtre commun qui leur est
propre et qui possédait déjà ce caractère.
Le groupe des primates est actuellement peu diversifié avec seulement 190 espèces. La plupart des grands singes
sont en voie d’extinction (territoires réduits, chasse ou braconnage). Néanmoins, les premiers primates fossiles
datent de 65 à -50 millions d'années ( Darwinius masillae Ida –47 Ma). Ces fossiles sont variés et ne sont
identiques ni à l'Homme actuel, ni aux autres singes actuels. La diversité des grands primates connue par
les fossiles permet de dire que ce groupe a été diversifié et que cette diversité est aujourd’hui réduite.
Au sein des primates, l’Homme appartient au groupe des Hominidés qui contient le Gorille, le Chimpanzé (Pan
troglodytes) et le bonobo (Pan paniscus).
Tous les hominidés sont caractérisés par différentes innovations évolutives : des narines rapprochées et une
absence de queue (remplacée par le coccyx). Ils possèdent notamment des modes de locomotion variés (bipédie
imparfaite, knuckle walking …) et des aptitudes à l’apprentissage et à l’empathie (attention envers ses
congénères).

2. La proximité entre le Chimpanzé et l’Homme
a. La phylogénie moléculaire
Lorsque les critères morpho- anatomiques ne sont plus suffisants, il est nécessaire de réaliser des phylogénies
moléculaires qui sont basées sur la comparaison des séquences nucléotidiques ou protéiques. La plupart des
séquences analysées montrent que l’Homme et le chimpanzé (+/- Bonobo) sont les espèces les plus proches des
Hominoïdes et partagent un ancêtre commun récent. Malgré le grand nombre de fossiles identifiés, aucun d’entre
eux ne peut être considéré à coup sûr comme un ancêtre de l'homme ou du chimpanzé.
Chez des espèces différentes, on peut rencontrer des molécules homologues. Il s’agit de protéines ou de gènes
dont les séquences montrent des ressemblances importantes, ne pouvant être attribuées au hasard. Les espèces
possédant ces molécules doivent avoir une origine commune. On utilise les homologies moléculaires pour préciser
le degré de parenté entre les espèces : des espèces sont d’autant plus apparentées que les séquences de leurs
molécules homologues sont semblables. En appliquant ce raisonnement aux molécules d’opsines contenues dans les
cônes (1S) ainsi qu’à d’autres molécules (comme le gène BRCA1 électrophorèse TP), on confirme que l’Homme fait
partie des primates, et on démontre qu’au sein des primates hominoïdes son plus proche parent est le chimpanzé
(Chimpanzé au sens large c’est-à-dire : chimpanzé commun et bonobo).
Hominoïdes

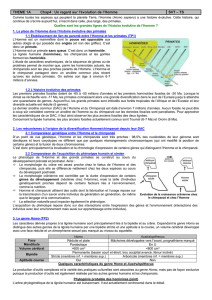
Matrice des pourcentages de différences entre séquences d’acides aminés de la cytochrome oxydase ( enzyme
impliquée dans le chaîne respiratoire mitochondriale) de différentes espèces de primates.
Arbre phylogénétique construit à partir de la matrice des distances pour la cytochrome oxydase de différentes
espèces de primates
b. Analyse des caryotypes
D'un point de vue génétique, l'Homme (46 chromosomes) et le chimpanzé (48 chromosome) sont très proches. Les
séquences nucléotidiques comportent seulement 1,5% de différences (mutations) soit seulement 10 fois plus
qu’entre deux individus humains pris au hasard. Les différences entre les caryotypes sont expliquées simplement
par la position de certains gènes : des remaniements chromosomiques (translocation, fusion), de phénomènes de
duplications géniques (p 27 enzyme AMY1 amylase : le gène de l’amylase est présent en plusieurs exemplaires
(duplication génique) chez l’Homme notamment chez l’individu d’une population ayant un régime alimentaire
riche en amidon (sélection naturelle)), de pertes de séquences génétiques et de mutations ponctuelles.
Homme
Chimpanzé
Gorille
Orang-Outan
Homme
0
Chimpanzé
1 fusion
0
Gorille
1 fusion
1 inversion
1 inversion
0
Orang-Outan
1 fusion
2 inversions
2 inversions
1 inversion
0
Tableau des remaniements chromosomiques entre les chromosomes 2p, 2q de 3 grands singes et 2 de l’Homme
c. La mise en place du phénotype humain ou simien
Néanmoins, ces proximités génétiques n’expliquent pas les nombreuses différences entre ces deux espèces. En
effet, le phénotype de l’Homme et du chimpanzé se construit au cours du développement pré et post- natal,
sous l’effet de différences dans la chronologie et l’intensité d’expression de mêmes gènes (hétérochronie)
en interaction avec l’environnement (dont la relation aux autres individus). Ainsi la morphologie du crâne des
fœtus humains et de chimpanzés est proche, mais au cours du développement pré et post natal, les variations de
l’expression de gènes homéotiques (s’exprimant à des moments et sur des périodes différentes au cours du
développement) modifient profondément le phénotype.

Ainsi chez Homo sapiens : crâne volumineux, face plane, mâchoire parabolique, absence de crocs, trou occipital
centré, colonne vertébrale avec quatre courbures, bassin court et large, fémurs convergents, membres inférieurs
plus longs que les membres supérieurs (chez le singe adulte mâchoires développées fortement au détriment de la
boite crânienne et ainsi modeste volume de l’encéphale (400-450 cm3). D’autre part, l’environnement joue un
rôle important dans le domaine de l’acquisition du langage (chapitre 2 p50/51 transmission culturelle des
comportements) ou des comportements comme la fabrication d’outil.
Homme seul animal à émettre des sons articulés grâce à un grand pharynx et à un larynx bas contrairement aux
singes.
Evolution du crâne de l’Homme et du
Chimpanzé lors du développement postnatal

Portrait hypothétique de l’ancêtre commun :
Possession de tous les caractères dérivés que se partagent l’homme et les chimpanzés, autour de -7Ma. Taille
moyenne de 1 m pour 30 à 40 kg. Encéphale assez développé. Faculté de se déplacer temporairement uniquement
grâce aux pattes de derrière. Vie essentiellement en milieu arboricole.
Bassins et fémurs de quelques grands primates
II. Quels sont les caractères propres à la lignée humaine et quelles interrogations pose la reconstitution
précise de l’arbre phylogénétique du genre Homo ? TP n°8
Lignée humaine : totalité de l’histoire évolutive des hominines à partir de l’ancêtre commun chimpanzé-homme
jusqu’à l’être humain actuel, environ 7Ma.
1. Les critères d’appartenance au genre Homo
Le genre Homo regroupe l'Homme actuel et quelques fossiles qui se caractérisent notamment par une face
réduite et un angle facial presque droit, une mandibule parabolique, un dimorphisme sexuel peu marqué sur
le squelette, un volume crânien important (1400 cm3), des adaptations à la bipédie permanente et à la
course à pied avec notamment un trou occipital centré sous le crâne, la présence de courbures sur la colonne
vertébrale, bassin avec des os iliaques courts et larges pour permettre l’insertion de puissants muscles
fessiers indispensables à la station debout, des pieds à orteils courts et pouce non opposable, des bras plus
courts et des fémurs convergents vers les genoux …
La production d'outils complexes et la variété des pratiques culturelles sont associées au genre Homo, mais
de façon non exclusive. La construction précise de l’arbre phylogénétique du genre Homo est controversée
dans le détail.
 6
7
8
6
7
8
1
/
8
100%