Thème 1A : GENETIQUE ET EVOLUTION Chapitre 1 : Le brassage

1
Thème 1A : GENETIQUE ET EVOLUTION
Chapitre 1 : Le brassage génétique et sa contribution à la diversité génétique
Au sein d’une espèce, une grande diversité de phénotypes est observable. Les mutations contribuent pour une part
à cette diversité en créant de nouveaux allèles. D’autre part, chez les organismes pluricellulaires, la reproduction
sexuée comportent des évènements fondamentaux impliqués dans la transmission des chromosomes et donc des
allèles, et ainsi participent à l’obtention d’une diversité de génotypes.
Comment la méiose et la fécondation, évènements clés de la reproduction sexuée, contribuent-elles chacune
à la diversité génétique des individus ?
I. Comment s’effectue la méiose soit la production de cellules haploïdes à partir d’une cellule diploïde ?
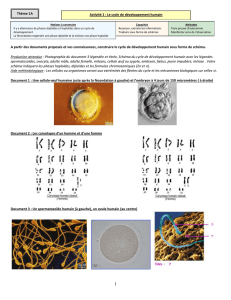
TP n°1
1. La stabilité du caryotype d’une espèce
Au cours du cycle de développement d’une espèce, on observe une alternance de phase diploïde des cellules
germinales et haploïde des gamètes. La méiose est le processus qui permet de produire de telles cellules haploïdes
à partir d’une cellule diploïde. Dans son schéma général, elle produit quatre cellules haploïdes à partir d’une cellule
diploïde et se déroule au niveau des cellules germinales (testicules, ovaires, étamines et ovaires des végétaux).
La fécondation, quant à elle, réunit deux cellules haploïdes pour former une cellule-œuf diploïde. Ces deux
mécanismes, méiose et fécondation, se succèdent au cours de tout cycle de développement et permettent (sauf
accident) la stabilité du caryotype d’une génération à la suivante.
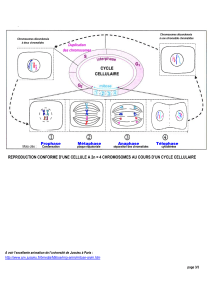
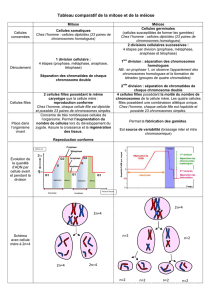
2. Les étapes de la méiose
La méiose est formée de 2 divisions cellulaires successives. Comme lors de la mitose, elle nécessite une étape
préalable de réplication de l’ADN (phase S Synthèse doublement de la quantité d’ADN). Chaque chromosome est
donc double c’est-à-dire constitué de deux chromatides identiques.
a. La première division de méiose (division réductionnelle).
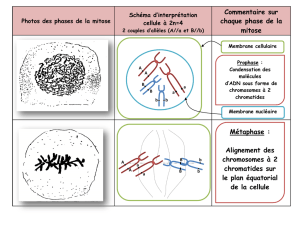
La première division de méiose permet la condensation et ainsi l’individualisation des chromosomes (prophase 1).
Les chromosomes homologues s’accolent sur toute leur longueur constituant ainsi n paire de chromosomes
homologues ou bivalents (appariement des K homologues déterminant). Ceux-ci vont ensuite se placer au niveau du

2
plan équatorial de la cellule (métaphase 1 : centromères de part et d’autre du plan équatorial). Sur chaque fibre
du fuseau de division se trouvent donc 2K homologues qui se font face. Tirés par les fibres du fuseau de division,
les 2K homologues de chaque paire se séparent (anaphase 1). Un lot haploïde de chromosomes doubles migre vers
un pôle de la cellule et un autre vers le pôle opposé. Deux cellules filles se forment par partage du cytoplasme de
la cellule mère, contenant chacune n chromosomes doubles à deux chromatides (télophase puis cytodiérèse).
C’est donc lors de cette première division que l’on passe de 2n chromosomes à 2 chromatides à n chromosomes à
2 chromatides. Cette première division est dite réductionnelle car elle diminue de moitié le nombre de
chromosomes.
b. La seconde division de méiose ou méiose II (division équationnelle).
La méiose II assure la séparation des chromatides de chaque chromosome. Les chromosomes subissent une
nouvelle prophase 2, métaphase 2 (centromères disposés sur le plan équatorial). Les chromatides sœurs des
chromosomes se séparent (anaphase 2 : clivage des centromères) et les K se répartissent dans les 4 cellules filles
(télophase 2 et cytodiérèse 2 : individualisation des cellules filles).
Ainsi, on passe de n chromosome à 2 chromatides à n chromosome à une chromatide. Cette division est dite
équationnelle car on conserve la formule n= x chromosomes, et se déroule comme une mitose mais sur des cellules
à n chromosomes.
La méiose, dans son schéma général, produit donc 4 cellules filles haploïdes à partir d’une cellule mère
diploïde. La méiose permet donc la production de gamètes ne contenant pas tous le même patrimoine génétique
(ex X ou Y).
II. Comment la méiose assure-t-elle en partie la diversité génétique des êtres vivants ? TP n°2
1. Définitions
Le monohybridisme est le suivi au cours des générations d’un caractère gouverné par un gène possédant 2 allèles.
Dans toutes les cellules, à l’exception des gamètes, chaque chromosome est présent en deux exemplaires portant
chacun un allèle d’un gène donné. Dans ce cas, les individus homozygotes produisent un seul type de gamètes alors
que les individus hétérozygotes produisent 2 types de gamètes.
Le croisement test. Voir fiche méthodologique génétique.
Afin de déterminer le génotype d’un individu (et les gamètes produits), il est important de réaliser un croisement
test (test cross). Ce croisement consiste à croiser un individu à tester hétérozygote (comme F1) pour un caractère
avec un individu homozygote récessif pour ce caractère.
Le dihybridisme est le suivi au cours de générations de 2 caractères gouvernés chacun par 1 gène possédant 2
allèles différents. Dans ce cas, le nombre de gamètes varie entre 1 (pour le double homozygote) et 4 (pour le
double hétérozygote).
2. Le brassage interchromosomique
En anaphase 1 de méiose, les chromosomes homologues bichromatidiens de chaque paire se séparent (disjonctions
des couples d’allèles) et migrent aléatoirement et indépendamment pour toutes les paires. L’analyse statistique
des génotypes des descendants donne 4 phénotypes attendus équiprobables 25%. C’est le brassage
interchromosomique (gènes indépendants). Voir schémas.
Les différentes informations d’un croisement sont recensées dans un échiquier de croisement avec :
Les différentes types de gamètes.
Les différents génotypes.
Les différents phénotypes (et fréquence).

3
Le nombre de gamètes différents correspond à 2n, n étant le nombre de paires de chromosomes. Un humain peut
donc produire 2 23 gamètes soit 8 388608 possibilités. Cette évaluation est en fait largement sous évaluée, car
ce brassage interKique s’applique en réalité à des chromosomes préalablement remaniés par le brassage
intraKique. Le nombre de gamètes génétiquement différents produits par un individu est donc beaucoup plus
important.
3. Le brassage intra-chromosomique (remaniement).
Lors de la prophase 1, les K homologues s’accolent (tétrades) et entrecroisent leurs chromatides (chiasma) :
appariement. Dans le cas de gènes liés (locus sur le même K), la position de certains allèles peut alors s’inverser.
Au niveau des chiasmas, des portions de chromatides peuvent alors s’échanger. Des combinaisons non parentales
d’allèles apparaissent sur une chromatide et les chromatides sœurs ne possèdent plus le même contenu génétique.
Ce processus est appelé crossing-over ou enjambement. Ce CO est non systématique et aléatoire pour un couple
de gènes liés, la fréquence de recombinaisons est d’autant plus élevée que la distance entre eux est grande sur le
K.
Dans ce cas, les 4 phénotypes sont présents dans la descendance avec 2 phénotypes parentaux surreprésentés et
2 phénotypes recombinés sous-représentés.
Si k nombre moyen de locus à l’état hétérozygote par chromosome, nombre de gamètes différents produits à
l’issue des processus méiotiques est : 2 nk
Et donc pour l’espèce humaine, k=100 soit 2 2300 !!!!!
La méiose, par ces doubles brassages chromosomiques, permet la production d’une diversité potentiellement
infinie de gamètes
BRASSAGE INTRACHROMOSOMIQUE
a
a
b
b
a’
a’
b’
b’
a
a’
a’
a
b
b
b’
b’
b
b
a
a’
b’
b’
a
a’
b
a
b
a’
b’
a
b’
a’

4
III. Comment la fécondation est-elle une source de diversité génétique supplémentaire ? livre p24/25
La fécondation se déroule de manière identique chez la plupart des animaux. Un gamète mâle (n) et un gamète
femelle (n) s’unissent. Généralement, l’entrée du spermatozoïde rend l’ovule « imperméable » aux autres
spermatozoïdes. Quelques heures plus tard, se produit la caryogamie : fusion des pronucléus (noyaux haploïdes)
reformant ainsi un noyau diploïde et ainsi constitution d’une cellule-œuf (zygote).
Lorsque que le contact s’effectue entre les deux membranes, l’ovocyte 2 achève sa méiose et devient ainsi l’ovule
pour un temps éphémère. Les noyaux du spermatozoïde et de l’ovule gonflent, ceci étant dû à une réplication de
leur ADN : ainsi les chromosomes à 1 chromatide repassent à 2 chromatides. Ils se transforment ainsi en
pronucléus mâle et femelle. Il y a rapprochement et fusion 15 à 20 heures après l’arrivée du spermatozoïde
(caryogamie). Il y a ainsi constitution de l’œuf ou zygote qui entame aussitôt sa division par mitoses.
Photo de caryogamie (MO)
Pronucléi indiqués par pointes flèches rouges
La diversité génétique potentielle des zygotes est immense car elle résulte de la rencontre aléatoire entre les
gamètes mâle et femelle. Ces gamètes sont issus d'une quasi-infinité de gamètes possibles possédant chacun une
combinaison d’allèles inédite pour les différents gènes du génome. La fécondation amplifie donc le brassage
génétique réalisé lors de la méiose et le nombre d’assortiments chromosomiques différents résultant de la
reproduction sexuée est ainsi quasiment infini.
Pour un couple humain, le nombre théorique d’enfants à génome différent : 2 2300 X 22300 !!!
Seule une fraction des zygotes est viable et se développe car le taux d’échec est important à chacune des étapes
du développement. De plus, des erreurs peuvent se produire.

5
IV. Quelles sont les différentes anomalies survenant au cours de la méiose, et quelles en sont les
conséquences sur la descendance ? TP n°3
1. L’origine des familles multigéniques.
La très grande majorité des CO correspondent normalement à des échanges de portions parfaitement homologues
de chromatides. Parfois, des appariements incorrects des chromosomes se produisent et ainsi des anomalies
peuvent survenir. Un CO inégal aboutit parfois à une duplication de gène. Une des chromatides possède une partie
de matériel génétique supplémentaire (en double) alors que l’autre en perd une partie. Ce mécanisme est à l’origine
de la duplication génique et, est ainsi source de diversification du vivant.
Prophase 1
Des cellules œuf
Des spermatozoïdes Des ovules
CROSSING OVER INEGAL
Les copies ainsi dupliquées (+ translocation soit insertion de la copie ailleurs dans le génome soit au niveau du
même chromosome ou non) peuvent subir des mutations et évoluer indépendamment. La quantité de mutations
s’accumule au cours du temps. Ainsi, plus les séquences des gènes sont éloignées, plus la duplication génique est
ancienne.
Ce phénomène contribue à la formation de nouveaux gènes. Il ne s’agit pas d’allèles du même gène : en effet, ces
séquences ont des locus différents. Ces gènes codent pour des protéines proches (mais qui peuvent avoir des rôles
différents) et restent très apparentés (entre 25 et 99% d’homologie). Ils forment une famille multigénique.
Les duplications, les transpositions et les mutations sont à l’origine de la diversification des génomes et des êtres
vivants. L’histoire évolutive des familles multigéniques peut être retracée par la réalisation d’un arbre
phylogénétique basé sur les séquences des gènes ou protéines étudiées. Seulement duplication-translocation de
gènes, mais sans mutation : obtention d’un grand nombre de copies identiques de gènes éparpillés dans le génome
(permet l’amplification de la synthèse de ces gènes : exemple amylase entre homme et chimpanzé p27).
B
B
A
A
B
B
A
A
B
A
A
A
B
B
A
B
A
B
A
A
B
A
A
B
A1
A
B
A
A2
B
A1
A2
B1
A1
A2
B
A1
A
B
A
A2
B
A
A
B
 6
6
1
/
6
100%