Télécharger le fichier - Fichier

2015-2016 La physiologie bactérienne
La physiologie bactérienne
– UEVII: BPV–
Le code génétique (suite)
Semaine : n°11 (du 16/11/15 au
22/11/15)
Date : 19/11/2015
Heure : de 9h00 à
10h00 Professeur : Pr. ROMOND
Binôme : n°D9 Correcteur : D8
Remarques du professeur : Aucune.
PLAN DU COURS
III) Conjugaison
A) Observation
1) Raisonnement
IV) La fertilité
A) Le facteur F et les caractéristiques de transfert.
1) Transfert de F+ vers F-
2) Transfert de HFR vers F-
3) Transfert de F' vers F-
4) Résumé des types transferts :
V) Les plasmides
A) Généralités
B) La pression antibiotique
VI) La transduction
A) Le cycle lithique
B) Le cycle lysogénique
1/9

2015-2016 La physiologie bactérienne
III) Conjugaison
On avait envisagé la conjugaison par les expériences de Lederberg et Tatum suivi par Devis sur le principe d'une
bactérie (Esch. Coli.) qui aurait eu des exigences différentes au seins d'une espèce.
Est-il nécessaire d'avoir un contact entre les bactéries ?
Devis a montré que pour qu'il y ait acquisition de critères qui permettaient sur un milieu minimum de cultiver, il
fallait un contact étroit car le phénomène du tube en U empêchait l'acquisition du transfert de gènes.
Si on met en contact les bactéries A et B avec un ensemencement, on arrive à avoir une colonie.
Est ce qu'il y a un sens d'échange de gènes ?
Dans le cas de Ledeberg et Tatum, deux souches d'Escheria Coli. avaient les mêmes potentialités de transfert.
Il faudrait disposer de souches complémentaires (mutantes) qui ont les critères de dépendance (méthionine..), on a
la première souche A que l'on transforme de façon à avoir deux souches :
–une sensible à la streptomycine (A Sensible)
–une autre résistante à la streptomycine (A Résistante)
Chaque souche complémentaire (A et B) aura deux sous classes (deux nouvelles souches) avec une Résistance (R)
et une Sensibilité (S) à la streptomycine (antibiotique de sélection).
Expérience de Lederberg et Tatum en associant :
–A Sensible avec B Sensible
–A Sensible avec B Résistante
–A Résistante avec B Sensible
–A Résistante avec B Résistante
Après un temps de contact de quelques heures, on ensemence une gélose contenant de la streptomycine comme
sélecteur, c'est un milieu minimum donc seules les bactéries qui ont réceptionné les gènes complémentaires vont
pouvoir cultiver.
On ensemence chaque mélange sur un milieu minimum avec le sélecteur.
A) Observation
On observe que lorsqu'il y a deux souches sensibles (S) ensemble, il n'y a pas de culture, cela veut dire que la
souche qui réceptionne les gènes est unique. Les deux ne vont pas pouvoir réceptionner les gènes. Si on a les
mêmes gènes présents avec un transfert dans les deux sens logiquement avec A (R) et B (R) on aurait eu une
culture, en fait il n'y a qu'avec A (R) et B (S) qu'il y aura une culture, c'est A qui réceptionne les gènes de B car
elle pourra cultiver.
A (R) ne peut pas transférer à B (S), B va résister et cultiver car le deuxième critère c'est le milieu minimum, il
faut réceptionner des gènes complémentaires à ceux déjà acquis.
Il y a bien un sens.
1) Raisonnement
J'ai deux exigences de culture pour cultiver sur milieu minimum :
Il nous faut des gènes indépendants de substrats (méthionine cultivé etc.)
Il faut une mutation par contact étroit pour apporter un complément de gènes.
Est ce que ça va dans les deux sens ?
2/9

2015-2016 La physiologie bactérienne
Non, car il y a un sélecteur à la streptomycine et si il y a résistance des deux côtés, on ne peut rien faire.
Il faut qu'il y ai résistance de la bactérie qui réceptionne pour cultiver, si on donne à A(S) par B(R) il n'y aura pas
les gènes protégeant du sélecteur.
Il y 2 systèmes de sélection :
–La caractéristique du milieu minimum : c'est un milieu où il n'y a pas les 4 composés, il nous faut une
bactérie autotrophe pour cultiver en absence de leucine etc.
–Résistance à un système de multiplication, le fait de ne pas avoir de culture dans les deux systèmes
montre qu'il y a une direction du transfert, les deux souches n'ont pas la même capacité bien
qu'appartenant à la même espèce.
Il y a un transfert dirigé, donc A et B ne sont pas équivalents dans la capacité à transférer les gènes, donc on a
extrait L’ADN pour connaître la spécificité.
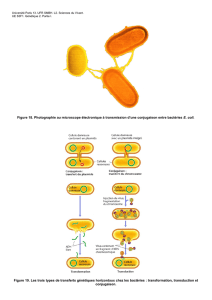
Quand on regarde, nous avons A récepteur et B donneur et lorsqu'on extrait de
l'ADN B et qu’on centrifuge en chlorure de césium (ce qui permet la migration
en fonction de la densité de l’ADN) on s'aperçoit que chez B il y a deux anneaux
correspondant à deux structures d’ADN double brin.
Si on observe A, il n'y aura que le chromosome avant mis en contact avec B.
Après mise en contact nous avions en plus du chromosome (correspondant à la
mémoire génétique de la souche) un élément de l’ADN double brin en position
extra chromosomique qui a pu être transféré.
IV) La fertilité
On a donc la sexualité des bactéries car à partir du moment où, dans une même espèce, il y a des individus qui
peuvent transférer à un autre individu des codes génétiques alors ils ne sont pas équivalents dans la fonctionnalité
(transfert de code), ici le premier tamis sur lequel on a travaillé c’est le facteur F dit de fertilité.
Ce facteur F a été le premier plasmide étudié qui nous a permis de classer l’ensemble des plasmides découverts.
A) Le facteur F et les caractéristiques de transfert.
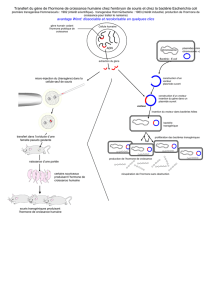
On va avoir plusieurs possibilités, soit le facteur F est extra chromosomique (F+ vers F-), à ce moment là, il faut
que la Bactérie B (modèle) soit en contact étroit avec la bactérie A.
Que se passe t-il au moment du contact ?
Dans le Facteur F, il y a le code pour les pilis (un pilus, des pilis) qui vont permettre une adhésion entre la
bactérie B porteuse du plasmide et la bactérie A (sans plasmide), cela forme une sorte de canal où pourra passer
l'ADN.
1) Transfert de F+ vers F-
F+: présence du plasmide extra-chromosomique (ADN circulaire)
F-: n'a pas de compétences génétiques suffisantes pour effectuer un transfert.
3/9

2015-2016 La physiologie bactérienne
Au moment où l'on crée le pont, il y a l'initiation de la réplication du plasmide.
Entre le plasmide et le chromosome, il y a une indépendance de réplication, il n'y a pas nécessairement de
réplication du chromosome.
On pourra démarrer la réplication du plasmide quand il n'y a pas de réplication du chromosome.
Deux phénomènes possibles : il y a donc la réplication bidirectionnelle pour le chromosome et la réplication en
cercle roulant qui intéresse le plasmide.
Il y a l’ouverture d’un seul brin, on aura alors un déroulement du simple brin qui sera poussé par le plasmide vers
la bactérie A.
Le système commence, le plasmide s'ouvre au niveau du point de réplication et le brin 5’ est poussé vers la
bactérie F- (A), le mécanisme de réplication se met en route de façon à avoir un plasmide lorsque la réplication est
complète.
Quand on a attendu le temps que tout le plasmide passe avec le monobrin (qui n'est pas parti de la
bactérie B) le brin est répliqué sur la bactérie B ainsi que dans la bactérie A et le pont disparaît.
On a une réplication double à l'intérieur de F+ et de F-. Le brin va être répliqué des 2 côtés.
La bactérie A qui était F- est devenue F+ et B est restée F+, on ne transfert pas en perdant le plasmide ; au contraire
on le conserve dans la première bactérie F+, on a donc un transfert élevé de plasmide.
Il y a une deuxième possibilité (pas la principale) : le plasmide peut se retrouver dans le chromosome, il sera donc
intra-chromosomique dit sous forme épisomique.
Cette forme épisomique s'appelle une forme HFR (Haute Fréquence de Recombinaison).
Il y a des séquences d'insertions dans le chromosome qui permettent qu'on ait l'ensemble du plasmide F qui soit
dans ce chromosome.
2) Transfert de HFR vers F-
Le plasmide inséré dans le chromosome n’a pas perdu ses capacités d’origines de réplication qui sont insérées à un
endroit donné du chromosome et non plus au point d'insertion et elles sont toujours indépendantes, même dans le
chromosome.
L'origine de la réplication est toujours indépendante de la réplication dans le chromosome, on a toujours le code
fonctionnel aux pilis.
Comment se met en place la conjugaison ?
Il faut un contact entre la bactérie HFR et la bactérie F- (Bact. A, dépourvue de plasmide). Sur HFR, le code est
toujours présent pour les pilis, accrochage à la bactérie F- etc.
Il y a un système en cercle roulant, il y a donc l’origine de la réplication, comme c'est un système en cercle
4/9

2015-2016 La physiologie bactérienne
roulant il y a un mono brin qui va s’ouvrir, on déroule le plasmide, ce qui nous donne F'.
La première étape consiste à faire un brin complémentaire, pour faire passer tout le plasmide de la bactérie HFR à
la bactérie F- → il faut beaucoup de temps.
Au final, il y a (avec le système en cercle roulant ayant répliqué) la partie plasmidique qui est peu présente et qui
reste en entier en double brin.
Sur le HFR, on n'a pas perdu le plasmide, mais une partie du plasmide et de l’ancien chromosome sont passées
vers le F- ce qui donne F'. Le chromosome de HFR ne sera donc pas totalement fonctionnel, et la partie de la
souche du chromosome HFR peut se recombiner avec le gène équivalent dans le chromosome de la bactérie F -. On
parle donc de Haute Fréquence de Recombinaison.
Il s'effectue une recombinaison entre la structure presque plasmidique et le chromosome, car même sur des
bactéries qui ont des différences de souches, celles-ci ont une partie de leurs génome en commun et quelques
petites différences au niveau de certains gènes.
3) Transfert de F' vers F-
Les souches F' ne pourront pas réinjecter des plasmides car cs derniers sont incorrects : ils ont perdu une partie
de leurs fonctionnalités donc la capacité de refaire le pilus.
Alors que la souche qui était HFR peut repasser en F+ et re-transférer le plasmide etc.
Le plasmide correspond au niveau des séquences génétiques à ses besoins : origine de réplication, insertion,
capacité de synthétiser le pilus et de pousser le plasmide vers une autre bactérie.
Le plasmide a un gène facilement transférable, car ré-acquérir des gènes du chromosomes est difficile mais
possible si on a une excision du plasmide HFR. Il peut y avoir des erreurs et au lieu de couper une zone
d’insertion, il y a une coupure en reprenant une partie du chromosome faisant perdre une partie du plasmide
donnant un plasmide F' ; si la zone d'erreur n'est pas trop importante il reste fonctionnel.
Le système F+ vers F- (extra chromosomique) : système facilement mobilisable et facilement transférable qui va
avoir une fréquence de recombinaison génétique faible et transmission à haute fréquence du facteur F.
Le plasmide sous forme épisomique (Hfr) HFR vers F- : le transfert du facteur F est faible mais la fréquence de
recombinaison génétique est élevée.
4) Résumé des types transferts
Lorsque l’on est dans le système F extra-chromosomique avec un transfert de F+ vers F-, une fois le contact :
–Synthèse des pilis.
–Ouverture du plasmide et la réplication sous forme de cercle roulant (copie qui va dans la cellule
receveuse et une autre qui reste dans la cellule émettrice).
–La receveuse ré-induit la réplication du plasmide, au final on a donc deux bactéries F+ mais avec une
faible fréquence de recombinaison et une haute fréquence de transfert du plasmide.
5/9
 6
7
8
9
6
7
8
9
1
/
9
100%