Cours de Neurophysiologie - FSA

SV5
LICENCE FONDAMENTALE : BIOLOGIE
CELLULAIRE ET MOLECULAIRE
Cours de Neurophysiologie
Pr. Ez-Zaher Latifa
Module
Endocrinologie/Neurobiologie
1

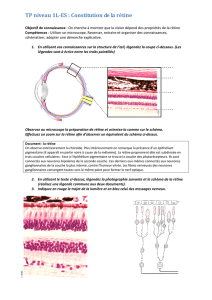
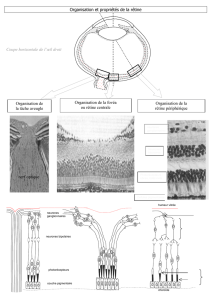
Composantes cellulaires du tissu nerveux
Structure du neurone
2

Le potentiel de repos est le résultat d’une différence de concentration en ions de part et d’autre de
la membrane et de la perméabilité sélective de la membrane cellulaire certains ions.
Milieu extracellulaire
Milieu intracellulaire
Concentration en millimoles / litre
Ions
Na+
15
150
45
400
15
150
45
400
K+
Cl-
Gros anions (A-)
3

Periode réfractaire
Pendant la période réfractaire
pour que
ire x
: un e
absolue l’inactivation est
maximale et la probabilité
les canaux K+ soient ouverts est
grande. Un second potentiel
d'action ne peut être émis.
Pendant la période réfracta
relative, l'inactivation des canau
Na+ et la probabilité pour que les
canaux K+ soient ouverts
diminuent progressivement
second potentiel d'action peut êtr
émis mais la valeur du potentiel
seuil est moins négative.
4

Fibre non myélinisée : Propagation continue
Fibre myélinisée saltatoireconduction
5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
1
/
45
100%