Bio Dev Cours 3

8
3. La spermatogenèse :
Le testicule assure deux fonctions :
o La production de cellules sexuelles mâles : c’est la spermatogenèse.
o La production d’hormones stéroïdes mâles : c’est la stéroïdogenèse. L’hormone la plus
importante est la testostérone.
Cf schéma p3
Ces deux fonctions sont associées à une compartimentation fonctionnelle du testicule. Il y a
tout d’abord les tubes séminifères, dans ces tubes a lieu la spermatogenèse. Ces tubes sont
disséminés dans un tissu interstitiel dans lequel sont localisé les cellules de Leidig responsables
de la production de testostérone.
Le testicule est composé de différents lobules. Au sein de ces lobules, on distingue les tubes
séminifères. Ces tubes se rejoignent/s’anastomosent au niveau d’un réseau, le Rete testis. Ce
réseau va déboucher dans l’épididyme.
L’épididyme est composé de tubes contournés qui vont s’étendre sur toute la longueur du
testicule. L’épididyme est composé de différente région : la tête, le corps et la queue. Il a un
rôle majeur dans le stockage mais surtout dans la maturation des spermatozoïdes. La maturation
des spermatozoïdes a lieu dans l’épididyme car lorsqu’ils sortent du testicule, ils ne sont pas
fécondants. Ils vont subir des remaniements membranaires caractérisés par des modifications
de lipides membranaires et résidus glucidiques. Ces remaniements vont faire apparaître un
système de reconnaissance qui va permettre de reconnaître l’ovocyte et plus particulièrement la
zone pellucide de l’ovocyte. Il y a également la réorganisation des différents organites, celles-ci
ont pour but de mettre à jour des récepteurs pour reconnaître l’ovocyte.
Il y a la fabrication d’un système de protection qui va recouvrir ces récepteurs afin de les
protéger durant tout le transit dans les voies génitales mâles et femelles.
Après être dans l’épididyme, les spermatozoïdes vont se retrouver dans le canal déférent.
Au sein des tubes séminifères, il y a la présence de différents types cellulaires qui peuvent être
très différents les unes des autres. La majeure partie de ces types cellulaires correspond à des
cellules qui sont à différents degrés de représentation de la spermatogenèse.
Au sein de ces tubes il y a donc des cellules somatiques et des cellules germinales.
On trouve des cellules somatiques très allongées ce sont des cellules péritubulaires ou cellules
myoïdes. Ces cellules entourent le tube séminifère, elles ont un caractère de cellules
musculaires (myoïdes). Ce caractère permet d’évacuer le fluide testiculaire et les
spermatozoïdes dans la lumière du tube séminifère.
A l’intérieur, on trouve les cellules de Sertoli, ce sont de grosses cellules avec un noyau pas
arrondi. Ces cellules envoient des prolongements cytoplasmiques qui entourent les cellules
germinales dans le tube séminifère. Les cellules de Sertoli ont un rôle majeur dans la
spermatogenèse : un rôle de soutien architectural, un rôle nourricier et un rôle différentiateur.
Il y a également différents types de cellules germinales. Plus les cellules germinales sont près
du pôle basal (au niveau des cellules myoïdes), moins ces cellules sont différenciées. Plus on se

9
trouve à l’intérieur du tube séminifère, plus elles sont différenciées. Ces différents types de
cellules germinales sont :
o Les spermatogonies de type A.
o Les spermatogonies de type B, qui sont déjà différenciées.
o Les spermatocytes à des degrés différents des étapes de méiose.
o Les spermatides, près de la lumière du tube séminifère, qui donneront les
spermatozoïdes.
1. Les différentes étapes de la spermatogenèse :
Cf schéma p3
La spermatogenèse est l’ensemble des processus qui aboutissent à la formation des
spermatozoïdes.
a) La phase de multiplication :
Cette phase correspond aux divisions goniales. Lors de cette étape, les spermatogonies A ont
deux voies possibles. Soit elles donnent de nouvelles spermatogonies A, soit elles rentrent en
différenciation pour donner des spermatogonies de type A, qui donneront par la suite les autres
types cellulaires. Cette étape à un double but : le 1er est de conserver le capital des cellules
souches en réserve, le 2nd est de multiplier le nombre de spermatozoïdes formés.
Chez l’homme, il y a environ 200millions de spermatozoïdes produit par jour, soit environ 2500
par secondes.
b) La méiose :
Cf schéma p4
La méiose débute à partir de la puberté, c’est un processus continu. Tout d’abord, les
spermatogonies de type A se différencient pour donner des spermatogonies de type B. on
rentre alors au stade de prophase de 1ère division. A la fin de la première division, on obtient des
spermatocytes II. On arrive alors en deuxième division au bout duquel on obtient des
spermatides.
NB : on observe les étape de méiose identiques à celles décrites dans le chapitre précédent.
c) La spermiogenèse :
Cf schéma p 4
La spermiogenèse concerne la transformation des spermatides en spermatozoïdes. Cette étape
permet la fabrication de cellules hautement spécialisées. Elle comprend une réorganisation du
noyau, la mise en place et le développement à partir de l’appareil de Golgi d’une structure : la
vésicule acromiale. Cette vésicule va se développée pour donner l’acrosome. Dans l’acrosome, on
trouve un sac rempli d’enzymes qui coiffent le noyau. Ces enzymes sont des enzymes
protéolytiques qui vont intervenir lors du franchissement de la zone pellucide par le

10
spermatozoïde, mais également dans l’assemblage des structures du flagelle. Pour cela, un des
deux centrioles va servir à la formation de l’axonème en flagelle.
En parallèle de cette formation, on observe une réorganisation des organites intracellulaires
notamment les mitochondries qui vont venir entourées la base du flagelle pour faire un manchon
mitochondrial. Celui-ci va apporter l’énergie nécessaire au mouvement du flagelle.
La maturation finale du spermatozoïde est entre autre d’acquérir une mobilité.
La dernière étape de la spermiogenèse est l’élimination des fragments cytoplasmiques. Ces
gouttelettes de cytoplasme sont appelées : corps résiduels. (Cellule rondecellule avec
tête+flagelle).
L’étape de spermiation correspond à la libération des spermatozoïdes dans
la lumière du tube.
Les cellules germinales présentent au cours de lors formation un certain nombre de caractère
qui détermine l’aspect histologique des tubes séminifères.
2. Caractéristiques spatio temporelles :
Cf schéma p5
La durée de la spermatogenèse est constante pour une espèce donnée (chez homme : 74 jours,
chez le rat : 48 jours), car chaque étape a une durée constante. L’entrée en spermatogenèse se
fait périodiquement tous les 16 jours chez l’homme.
Ces spermatogonies A entrent en groupe en spermatogenèse et reste reliées entre elles par des
ponts cytoplasmiques qui vont assurer le synchronisme de l’évolution du groupe. Comme la durée
de l’intervalle est régulière en un point du tube séminifère, on va avoir plusieurs cycle de
spermatogonies en spermatogenèse. L’évolution temporelle et spatiale de la spermatogenèse fait
que l’on peut observer 4 à 5 générations de cellules à un même moment dans le tube : c’est le
cycle de l’épithélium séminifère.
Ces générations de cellules germinales superposées forment des associations cellulaires
caractéristiques : ce sont les stades de l’épithélium séminifère. La succession de tous ces
stades correspond au cycle de l’épithélium séminifère.
4. L’ovogenèse :
L’ovogenèse se produit dans l’ovaire et a une triple fonction :
o La production de stéroïdes sexuels qui sont les œstrogènes et la progestérone.
o La production de cellules sexuelles, ces cellules sont les ovocytes = fonction gamétogene.
o Production des hormones permettant l’implantation et le développement de la gestation.
1. Introduction : l’ovocyte et le follicule :
Cf schéma p6
L’ovocyte lorsqu’il est produit est une grosse cellule qui devra être capable de fusionner avec un
spermatozoïde, de transformer son noyau et d’assurer les premières divisions de l’œuf. Pour

11
devenir apte à remplir ces fonctions, l’ovocyte doit subir une longue évolution cytologique,
moléculaire et fonctionnelle : c’est l’ovogenèse.
L’ovocyte est entouré par des cellules folliculaires, cette étape de croissance et de
différenciation et dépendante des cellules folliculaires qui entourent l’ovocyte : c’est la
folliculogenèse.
Le follicule est un ensemble cellulaire autour de l’ovocyte. L’ensemble des étapes qui aboutissent
à la formation d’un ovocyte mature est la folliculogenèse. Cette étape concerne une maturation
importante des cellules entourant l’ovocyte.
2. Les étapes de l’ovogenèse et de la folliculogenèse :
Tout débute au stade fœtal avec l’entrée en méiose et le blocage au stade diplotène. Lorsque la
puberté est atteinte, à chaque cycle juste avant l’ovulation, la méiose va reprendre pour donner
un ovocyte 2 qui va être bloqué en métaphase de division II de méiose. S’il y a fécondation, alors
la méiose reprendra et il y aura l’expulsion du 2ème globule polaire.
L’ovaire présente une structure très dense avec beaucoup de follicules. A la périphérie de
l’ovaire, on trouve les follicules primordiaux. Jusqu’à la puberté, des follicules vont se former, le
nombre de couches et donc de cellules entourant l’ovocyte va augmenter, pour former ainsi
différents stades de folliculogenèse. Jusqu’à la puberté, aucun follicule n’aboutira, on aura
toujours une dégénérescence.
Même après la puberté, la très grande majorité des follicules en maturation vont dégénérer : ce
phénomène est appelé atrésie folliculaire
(cf schéma p7).
Chez la femme, 70 à 99% des
follicules vont se dégénérer par atrésie.
L’entrée en croissance des follicules est un phénomène continu mais cela ne pourra aboutir avant
la puberté.
La maturation des ovocytes et des cellules folliculaires correspond à la folliculogenèse.
3. Les différents stades de l’évolution des follicules :
Cf schéma p7
On observe une phase de multiplication importante. Les follicules primordiaux (environ
40microns) vont évolué jusqu’à un follicule énorme (1 à 30mm). La multiplication cellulaire
intense permet la croissance du follicule.
Le follicule primaire (deux couches) puis les follicules secondaires et tertiaires eux ont plus de
couches qui les entourent. Toutes ces cellules qui entourent (l’ovocyte) sont des cellules de la
granulosa.
On observe également, un autre type cellulaire : les cellules de la thèque.
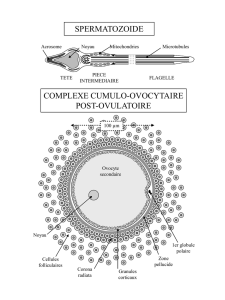
Les cellules qui entourent directement l’ovocyte forment la corona radiata (ce sont des cellules
de la granulosa).
La thèque est composée d’une partie interne et d’une partie externe.

12
Entre les cellules de la granulosa et les cellules de la thèque, on observe une membrane basale.
Et entre l’ovocyte et les cellules de la granulosa, on distingue également une membrane : c’est la
zone pellucide.
Les cellules vont se multiplier, la zone pellucide va se former entre la fin des follicules primaires
et le début des follicules secondaires.
La zone pellucide est une zone perméable aux molécules. Les cellules périovocytaires (cellules de
la granulosa) envoient des prolongements cytoplasmiques qui s’ancrent à l’ovocyte.
Ces jonctions qui relient les ovocytes aux cellules de la granulosa sont des jonctions perméables
assurant un couplage métabolique parfait entre la membrane plasmique de l’ovocyte et les
cellules folliculaires. Ce couplage permet des échanges permanent entre deux types cellulaires
et permet donc une synchronisation de la croissance ovocytaire et folliculaire.
Entre les différentes cellules de la granulosa, il y a aussi des jonctions communicantes. Au cours
de la folliculogenèse, il va apparaître au stade tertiaire une cavité : l’antrum. L’antrum va se
remplir de liquide folliculaire, va grossir et va finalement entourer l’ovocyte et la petite zone de
cellule de la granulosa. Cette petite zone correspond à la région du cumulus ophorius.
Au cours de cette croissance, il y a une multiplication cellulaire et une différenciation. Cette
différenciation sera une différenciation fonctionnelle.
Les cellules somatiques synthétisent les stéroïdes ovariens. Il y a une collaboration entre les
cellules de la granulosa et les cellules de la thèque pour qu’il y ait une synthèse d’œstrogène.
Il y a alors l’acquisition d’une capacité à répondre à l’hormone gonadotrope de type LH.
Au début de leur développement, les seules cellules possédant les récepteurs à la LH sont les
cellules de la thèque. Les cellules de la granulosa possèdent quant à elles, des récepteur à
l’hormone gonadotrope : la FSH. De plus en plus au cours de la croissance, on constate que les
cellules de la granulosa acquièrent des récepteurs à la LH, c’est l’étape finale. L’étape ultime
consiste à expulser l’ovocyte sous l’effet du pic de LH.
4. La croissance folliculaire : dynamique et régulation :
Cf schéma p8
a) L’initiation et la croissance basale :
Dans la 1ère partie, on distingue les follicules primordiaux, primaires et secondaires. Cette phase
est indépendante des hormones gonadotropes, mais elle dépende des facteurs de croissance
présents localement. Ces facteurs sont produits par les cellules de la granulosa et par l’ovocyte
lui-même. L’ovocyte produit la GDF9 qui intervient dans la différenciation et la multiplication
des cellules de la granulosa. Cette phase se nomme la croissance basale.
Les follicules primordiaux entament leur croissance basale pour devenir alors des follicules
primaires puis des follicules secondaires. Ces étapes sont indépendantes.
b) Le développement folliculaire terminal :
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%