Lire l`article complet

Mise au point
Mise au point
La Lettre du Neurologue - Vol. X - n°10 - décembre 2006
333
Mémoire et connectivité
Memory and connectivity
$G. Kalpouzos*, G. Chételat*, F. Eustache*, B. Desgranges*
* INSERM-EPHE-université de Caen, unité E0218, GIP Cyceron, CHU Côte-de-Nacre, Caen.
POINTS FORTS
Les interactions régionales cérébrales qui sous-tendent les
processus cognitifs suscitent depuis longtemps un intérêt
d’un point de vue fondamental mais aussi clinique ; néan-
moins, les dicultés techniques et méthodologiques n’ont
permis l’étude de la connectivité que très récemment.
Le fonctionnement de la mémoire épisodique dépend de
l’intégrité des structures limbiques et frontales, mais aussi
de l’état des bres de la substance blanche, qui connectent
ces structures entre elles.
Les diérentes régions entrant en interaction lors d’une
tâche de mémoire ne sont pas les mêmes selon le type de
traitement à eectuer, la nature du matériel à mémoriser ou
la population étudiée.
L’association de plusieurs techniques et approches
méthodologiques est nécessaire à la compréhension du fonc
-
tionnement cérébral en réseau. L’IRM fonctionnelle permet
de mettre en évidence les régions interagissant entre elles
lors d’une tâche cognitive, tandis que l’IRM du tenseur de
diusion constitue actuellement la seule technique in vivo
permettant la détection de microlésions au sein des faisceaux
de la substance blanche qui relient ces régions entre elles.
Mots-clés : Mémoire – Connectivité – Activation – IRM du
tenseur de diusion – Maladie d’Alzheimer – Hippocampe –
Cortex préfrontal.
SUMMARY
Memory is one of the cognitive functions that is the
most signicantly altered in diverse pathologies such as
Alzheimer’s disease.
Its functioning depends not only on the integrity of limbic
and prefrontal cerebral regions, but also on white matter
ber tracts which link these regions together.
Studying the multiple circuits that are recruited in memory
tasks is recent.
Currently, specic methodologies using data obtained with
functional magnetic resonance imaging (MRI) make it pos-
sible to highlight interactions between brain areas during
specic cognitive tasks, while data obtained with diusion
tensor imaging (DTI) provides accurate information about
the integrity of white matter bundles.
Keywords: Memory – Connectivit y – Activation – Diffusion
tensor imaging – Alzheimer’s disease – Hippocampus –
Prefrontal cortex.
W
ernicke (1874) bouleversa les conceptions structura-
listes de l’époque consistant à assigner une fonction
cognitive à une région cérébrale donnée. Sa théorie
associationniste postulait que les fonctions mentales supérieures
étaient le produit de connexions entre des aires corticales qui
traitaient des fonctions plus primaires. Ainsi, il estimait qu’un
trouble cognitif résultait d’une dysconnexion au sein de la subs-
tance blanche (pour une revue historique sur les syndromes de
dysconnexion, voir [1]). Chaque fonction mentale supérieure
dépend de structures spécifiques, mais ces régions ne peuvent
remplir leur rôle que si leurs connexions sont intactes, et cela
est particulièrement vrai pour la mémoire qui repose sur un
ensemble de structures cérébrales organisées en réseau.
Différents travaux en imagerie cérébrale utilisant des méthodo-
logies de plus en plus précises au sein de diverses populations
ont favorisé la mise en évidence des interactions régionales qui
sous-tendent les processus mnésiques. Ces travaux ont aussi
suggéré que des mécanismes de dysconnexion contribuent aux
troubles de la mémoire dans les pathologies neurodégénératives
comme la maladie d’Alzheimer (MA).
TROUBLES MNÉSIQUES DANS LA MA :
UN SYNDROME DE DYSCONNEXION ?
Il est établi que l’atrophie et le dysfonctionnement des régions
limbiques sont en grande partie responsables des troubles
mnésiques dans la MA. En effet, les études neuropatho-
logiques montrent une prédominance de plaques séniles et de

Mise au point
Mise au point
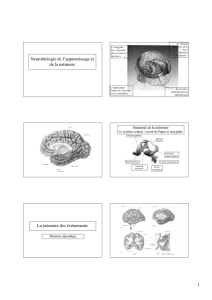
Figure 1.
Inuence de l’atrophie de l’hippocampe sur le dysfonc-
tionnement du cortex cingulaire postérieur chez le patient MCI
et dans la maladie d’Alzheimer.
Cingulaire
postérieur
Hippocampe
La Lettre du Neurologue - Vol. X - n°10 - décembre 2006
334
dont l’âge d’apparition des premiers troubles excédait les 65 ans.
En étudiant les corrélations entre les valeurs de métabolisme
de régions frontales (aires 10 et 9/46) et du précuneus (aire 7)
chez ces patients, nous avons pu montrer qu’une dysconnexion
fronto-pariétale serait en partie responsable de ces troubles
accrus. En effet, les corrélations positives étaient très signifi-
catives chez les sujets témoins, à la limite de la significativité
chez les patients à début tardif, et non significatives chez ceux
à début précoce.
Ces diverses observations corroborent l’hypothèse d’un syndrome
de dysconnexion dans la MA, qui contribuerait aux troubles de
la mémoire épisodique et de travail.
CONNECTIVITÉ FONCTIONNELLE DE LA MÉMOIRE
ÉPISODIQUE: ÉTUDES D’ACTIVATION
Sur la base d’études lésionnelles, Warrington et Weiskrantz
ont proposé en 1982 que le syndrome amnésique pouvait être
dû à une interruption des circuits neuraux spécifiques reliant
le cortex préfrontal au lobe temporal interne. Plus récem-
ment, les études d’activation ont montré que les processus
d’encodage et de récupération en mémoire épisodique étaient
sous-tendus par les régions du lobe temporal interne et du
néocortex, notamment des aires préfrontales, mais le rôle
spécifique de ces régions dans la mémoire a souvent été étudié
de façon indépendante. La figure 2 (A et B) résume les inter-
actions fonctionnelles entre ces régions lors de l’encodage et
de la récupération de l’information en mémoire épisodique
(5). Néanmoins, de nombreux facteurs sont susceptibles de
moduler ces interactions.
Eet de la profondeur du traitement du matériel
à mémoriser à l’encodage lors d’une tâche de
récupération
McIntosh et al. (6) ont mis en avant une connectivité différente
entre le cortex préfrontal et les structures du lobe temporal
interne en fonction de la condition d’encodage (traitement
superficiel ou profond des items). Indépendamment du type
de traitement à l’encodage, l’aire 10 du cortex préfrontal droit et
l’hippocampe gauche présentaient une connectivité fonctionnelle
constante lors de la récupération ; en revanche, l’aire 45/47 du
cortex préfrontal droit n’interagissait avec ces régions que lors
de la récupération des items encodés de façon profonde.
Eet de l’ecience de l’encodage sur le pattern
de connectivité fronto-hippocampique
Ranganath et al. (7) ont cherché à montrer les connexions
cortico-hippocampiques mises en jeu lors d’une tâche d’enco-
dage en mémoire épisodique, en distinguant les items encodés
avec succès (récupérés ultérieurement) des items dont l’enco-
dage a été inefficient (oubliés lors de la récupération). Ainsi,
les auteurs ont pu mettre en évidence l’évolution au cours du
temps des patterns de connectivité lors de la formation à long
terme de la trace mnésique. Les régions dont la connectivité
dégénérescences neurofibrillaires dans le lobe temporal interne
dès les premiers stades de la maladie. En ce qui concerne les
patients MCI (Mild Cognitive Impairment) chez lesquels les
lésions sont déjà étendues dans la formation hippocampique,
il existe un hypométabolisme de la région cingulaire posté-
rieure, elle-même peu touchée par ces lésions. Ces observations
récurrentes ont donné lieu à une hypothèse de dysconnexion,
via un phénomène de diaschisis, suggérant un effet à distance
de l’atrophie hippocampique sur le fonctionnement de base
du cortex cingulaire postérieur. Celui-ci est mis en évidence,
notamment sous forme d’un hypométabolisme en tomographie
par émission de positons (figure 1) [2]. Le dysfonctionnement
de ces deux régions clés pour la mémoire épisodique a aussi
été révélé au sein d’un réseau neuronal plus vaste par Greicius
et al. (3). Ces auteurs ont initialement mis en avant l’existence
d’un “mode de fonctionnement cérébral par défaut” sous-tendu
par un réseau complexe connectant diverses régions entre elles
et comportant notamment l’hippocampe et le cortex cingulaire
postérieur. Ce réseau a été mis en évidence en IRM fonctionnelle
(IRMf) au repos à l’aide d’une analyse des coactivations qui a
permis de montrer les régions dont l’activité varie ensemble lors
du repos. Ce circuit serait constamment activé à l’état de base,
mais son activation serait notablement réduite lors de tâches
faisant intervenir le traitement de stimuli externes. Dans un
groupe de patients atteints de MA où le dysfonctionnement
de ces deux régions a été confirmé, les auteurs ont conclu à
une altération globale de ce réseau qui pourrait être en partie
responsable de la dégradation de la mémoire épisodique.
Les lésions touchent aussi les régions associatives où siègent de
longues fibres corticocorticales, provoquant des déafférentations
au fur et à mesure que la MA progresse. Celles-ci contribue-
raient aux troubles mnésiques spécifiques, comme nous l’avons
récemment montré au sein de notre laboratoire (4). Ce travail a
mis en évidence une moindre efficience de la mémoire de travail
chez des patients à début précoce de la MA, comparés à ceux

Mise au point
Mise au point
Figure 2.
Interactions entre le cortex préfrontal et le lobe temporal interne lors des processus d’encodage (A) et de récupération (B) en mémoire
épisodique (schémas inspirés de Simons et Spiers, 2003).
A.
Encodage.
Selon ce modèle, (1) l’information à encoder en mémoire à long terme
est d’abord traitée par les aires corticales uni- et polymodales avant
d’être transférée au niveau du lobe temporal interne (LTI). Ce traitement
permet d’élaborer l’information perçue en une représentation uniée
composée de traits associés entre eux. (2) À ce stade, les interactions
avec le cortex préfrontal permettent un contrôle top-down des processus
d’encodage en fonction des objectifs à atteindre et de la demande de
la tâche. Elles garantissent le stockage à long terme de la représenta-
tion. (2a) Les régions préfrontales ventrolatérales sont engagées si les
processus requis sont de nature sémantique, la latéralité variant selon la
nature du matériel. (2b) Le cortex préfrontal dorsolatéral interagit si des
processus de sélection, de manipulation et d’organisation du matériel
à encoder sont requis.
B.
Récupération.
Selon ce modèle, (1) l’élaboration d’un indice de récupération en mémoire
dépend du cortex préfrontal ventrolatéral (avec une latéralisation variable
selon la nature du matériel) qui permet au LTI d’eectuer des comparaisons
et d’établir une concordance entre cet indice et l’information stockée en
mémoire.
Les informations plausibles réactivées en mémoire à long
terme sont maintenues en mémoire de travail grâce au cortex pré-
frontal ventrolatéral. (2) Alors, le cortex préfrontal dorsolatéral engage
des processus de contrôle an de sélectionner la bonne information
parmi toutes les autres. (3) Les aires préfrontales les plus antérieures
interagissent avec les autres régions si des processus cognitifs plus
complexes doivent être engagés, par exemple dans le cas d’une
génération interne d’informations. (4) Le cortex préfrontal médian
interviendrait spéciquement dans le cadre de la récupération de
souvenirs de nature autobiographique.
Régions postérieures
Renforcement des représentations
des informations perçues
Lobe temporal interne
Association des diérents éléments
constitutifs du souvenir
Cortex préfrontal ventrolatéral
Traitement sémantique et phonologique
de l’information à encoder
Cortex préfrontal dorsolatéral
Manipulation de l’information à encoder
Cortex préfrontal dorsolatéral
Manipulation et vérification
des informations récupérées
Cortex préfrontal antérieur
Contrôle mnésique de haut niveau
Lobe temporal interne
Comparaison entre l’indice
de récupération
et l’information stockée
en mémoire
Cortex préfrontal ventrolatéral
Élaboration d’un indice de récupération
Maintien de l’information retrouvée
Cortex préfrontal
médian 43
B
La Lettre du Neurologue - Vol. X - n°10 - décembre 2006
335
avec l’hippocampe gauche s’accroît lorsque l’encodage est réussi,
par rapport à un encodage inefficient sont :
le précuneus et les régions limbiques et paralimbiques dès
la présentation de l’item;
les cortex périrhinal, cingulaire postérieur, orbitofrontal
(aire 11) et frontopolaire ventral (aire 10) en phase précoce de
la formation de la trace;
le cortex frontal supérieur droit en phase tardive.
Cette étude montre que des régions distinctes participent à la
formation d’un souvenir au cours du temps, leur contribution
relative pour la réussite de l’encodage dépendant de l’intensité
des interactions avec l’hippocampe.
Eet de la composante émotionnelle
Il existe une amélioration des performances lorsque le matériel
à mémoriser est de nature émotionnelle. Quelques travaux en
neuro-imagerie suggèrent un rôle modulateur de l’amygdale sur
les régions du lobe temporal interne, ces régions interagissant
fortement entre elles. L’étude de Kilpatrick et Cahill (8) consis-
tait en une tâche d’encodage incident de films contenant des
éléments neutres ou négatifs, l’activité cérébrale étant mesurée
en TEP-FDG. Les résultats montrent notamment une forte
augmentation de l’influence directe de l’amygdale sur le gyrus
parahippocampique et le cortex orbitofrontal dans la condition
émotionnelle négative par rapport à la condition neutre.
Eet de l’âge
Des études menées auprès de sujets sains jeunes et âgés ont
permis d’apprécier les modifications des connexions survenant
au cours du vieillissement cérébral. L’étude de Daselaar et al. (9)
consistait en une tâche de récupération en mémoire épisodique
en IRMf. Les sujets âgés avaient tendance à reconnaître les items
davantage par sentiment de familiarité (réponses de type K
[know]) que par recollection consciente de l’information avec
son contexte d’acquisition (réponses de type R [remember]).
Par ailleurs, les réponses K dépendaient plutôt du cortex rhinal,
tandis que celles de type R dépendaient plutôt de l’hippocampe.
Les auteurs ont étudié les connexions fonctionnelles du cortex
rhinal d’une part et de l’hippocampe d’autre part, en fonction
de la réponse donnée par les sujets. Comparés aux sujets jeunes,
les sujets âgés présentaient une connectivité plus importante
entre le cortex rhinal et les régions préfrontales. Comparés aux
sujets âgés, les sujets jeunes présentaient une connectivité accrue
entre l’hippocampe et les cortex rétrosplénial (petite partie
du cortex cingulaire postérieur) et temporo-pariétal gauches.
D’après les auteurs, cette modification liée à l’âge reflèterait
un mode de compensation: la voie neuronale de la recollec-
tion étant altérée dans le vieillissement normal, les sujets âgés
recourent davantage à la voie neuronale de la reconnaissance
par sentiment de familiarité via les interactions entre le cortex
rhinal et les régions préfrontales.
A

Mise au point
Mise au point
Figure 3.
Organisation anatomo-fonctionnelle du circuit
de Papez (1937).
Néocortex
(aérences)
Faisceau
mamillo-thalamique
de Vicq d'Azyr
Gyrus cingulaire
Hippocampe Cortex entorhinal
Corps mamillaire
septum
Noyau antérieur
du thalamus
Aires sensorielles
polymodales
(projections)
Voie
Perforante
Piliers
du fornix
Capsule interne Cingulum
La Lettre du Neurologue - Vol. X - n°10 - décembre 2006
336
L’analyse de la connectivité effectuée à partir de données d’acti-
vation, bien que fondamentale du point de vue des interactions
entre les régions, présente certaines limites:
il est difficile de décider de l’orientation spatiale principale de
la connexion entre deux régions, et de savoir si une connexion
entre deux régions est de nature directe ou indirecte ;
au sein de populations différentes, il est difficile de conclure si
la connectivité différente est due à un changement de stratégie
ou à une réorganisation anatomique des réseaux.
Pour tester cette dernière hypothèse, il faudrait pouvoir analyser
finement l’intégrité des fibres de substance blanche, garante des
connexions régionales.
CONNECTIVITÉ ANATOMIQUE:
IRM DU TENSEUR DE DIFFUSION
Seule l’IRM du tenseur de diffusion (DTI) offre aujourd’hui la
possibilité de repérer, au sein de la substance blanche d’appa-
rence normale, des microlésions qui peuvent interrompre des
faisceaux de substance blanche. Cette technique repose sur
la détection des mouvements microscopiques des molécules
d’eau au sein des tissus. Elle fournit des index de l’intégrité de la
microstructure à travers la quantification de la “directionnalité”
(l’orientation spatiale principale) et du degré de diffusion de ces
molécules d’eau en chaque point du cerveau (respectivement,
l’anisotropie et la diffusivité). Ces mesures permettent d’aboutir
à la construction d’une cartographie des différents faisceaux
composant la substance blanche (tractographie). La maladie
d’Alzheimer provoque une démyélinisation, une perte axonale
et une augmentation de l’espace extracellulaire. Ces lésions
entraînent une diminution de l’anisotropie et une augmenta-
tion de la diffusivité dans la substance blanche qui connecte les
cortex associatifs (corps calleux, substance blanche des lobes
temporaux, frontaux et pariétaux), et au niveau de la substance
blanche du système limbique. Seule la technique de DTI permet
l’investigation des faisceaux spécifiques et de petite taille in
vivo, d’où son intérêt dans l’étude des processus mnésiques, qui
dépendent de l’intégrité de tels faisceaux comme le cingulum, la
voie perforante et le fornix (figure 3). L’intégrité du cingulum,
faisceau reliant le gyrus cingulaire postérieur aux structures
hippocampiques, a été testée par Fellgiebel et al. (10) chez des
patients à un stade prodromique de la MA. Ces auteurs ont
également associé la DTI et la neuropsychologie en corrélant
les indices d’anisotropie et de diffusivité à des scores mnési-
ques. Pour la première fois, une diminution de l’anisotropie
a été constatée chez les patients MCI par rapport à des sujets
contrôles, attestant de la détérioration de ces fibres de subs-
tance blanche. De plus, les auteurs ont montré une corrélation
significative entre les coefficients d’anisotropie et de diffusivité
du cingulum et les scores obtenus à une tâche de rappel verbal
différé, indiquant que cette dysconnexion entre deux structures
clés de la mémoire épisodique est en partie responsable des
troubles mnésiques. Chez des patients MCI et Alzheimer, Kalus
et al. (11) ont étudié l’intégrité de la voie perforante qui relie
le cortex entorhinal à l’hippocampe. Ce faisceau de fibres est
particulièrement vulnérable à la pathologie, puisqu’il se situe
dans la région où se manifestent les premières lésions. La dégra-
dation précoce de cette principale voie d’entrée dans le système
limbique pourrait être responsable des troubles précoces de la
mémoire. Le volume et l’indice d’anisotropie de trois régions
d’intérêt centrées sur l’hippocampe, le cortex entorhinal et la
voie perforante ont été mesurés. La seule différence signifi-
cative entre le groupe MCI et le groupe contrôle se situe au
niveau de l’indice d’anisotropie de la voie perforante. De plus,
l’indice d’anisotropie de la voie perforante, mais non pas son
volume, est corrélé aux scores de rappel différé, démontrant
la capacité de la DTI à détecter les dysconnexions en partie
responsables des déficits mnésiques au stade prodromique de
la MA. Cette technique, couplée à une analyse volumétrique, a
permis à Kuroki et al. (12) de tester l’intégrité du fornix, faisceau
reliant l’hippocampe aux corps mamillaires et au septum, chez
des patients atteints de schizophrénie présentant des troubles
mnésiques. Là encore, la DTI s’est révélée efficace pour mettre
en évidence des anomalies au niveau du fornix (diminution de
l’anisotropie et augmentation de la diffusivité), ces anomalies
étant de plus corrélées avec la diminution du volume hippocam-
pique. Néanmoins, seule cette dernière mesure était corrélée
aux scores mnésiques des patients.
Ainsi, chaque méthode prise isolément ne permet pas d’appré-
hender l’ensemble des modifications cérébrales survenant avec
l’âge ou la pathologie; le couplage de plusieurs techniques semble
nécessaire, comme l’atteste l’étude très récente de Persson et
al. (13). Ces auteurs ont étudié les corrélations structuro-fonc-
tionnelles du déclin mnésique au cours du vieillissement. Ils
ont ainsi montré en DTI l’altération préférentielle des fibres de
la région antérieure du corps calleux chez les sujets présentant
une dégradation plus importante de la mémoire épisodique, cela
associé à une augmentation d’activité spécifique de la région

Mise au point
Mise au point
AUTO-ÉVALUATION
I. Une dysconnexion spécique pourrait être en partie respon-
sable des troubles précoces de la mémoire épisodique dans la
maladie d’Alzheimer. Quelles régions cérébrales cette dyscon-
nexion concerne-t-elle ?
a. le cortex frontal et l’hippocampe
b. l’hippocampe et le cortex cingulaire postérieur
c. l’hippocampe et le thalamus
d. l’amygdale et l’hippocampe
II. Quelle technique d’imagerie cérébrale permet d’étudier direc-
tement et in vivo les faisceaux de substance blanche ?
a. TEP (Tomographie par émission de positons)
b. IRM fonctionnelle (Imagerie par résonance magnétique)
c. DTI (Imagerie du tenseur de diusion)
d. EEG (Électroencéphalographie)
III. De nombreuses régions cérébrales sont impliquées dans
les processus d’encodage et de récupération de l’information
en mémoire épisodique. Parmi ces propositions, laquelle est
fausse ?
a. le cortex préfrontal dorsolatéral et les régions occipitales
b. le cortex orbitofrontal et l’hippocampe
c. le cortex préfrontal et les structures striatales
d. les structures limbiques et le cortex préfrontal
Résultats : I : b ; II : c; III : c.
La Lettre du Neurologue - Vol. X - n°10 - décembre 2006
337
ventrale droite du cortex frontal (mesurée en IRMf), suggérant
que l’altération structurale du cerveau entraîne la mise en place
de processus compensatoires.
CONCLUSION
Actuellement, très peu d’études ont associé différentes tech-
niques d’imagerie cérébrale et la neuropsychologie, qui sont
pourtant complémentaires. Néanmoins, les travaux les plus
récents ayant couplé des données de natures différentes ont
permis une meilleure analyse des processus complexes de la
mémoire.
RÉFÉRENCES BIBLIOGRAPHIQUES
1. Catani OM, Ffytche DH. e rises and falls of disconnection syndromes. Brain
2005;128:2224-39.
2. Chételat G, Desgranges B, de la Sayette V et al. Dissociating atrophy and
hypometabolism impact on episodic memory in mild cognitive impairment.
Brain 2003;126:1955-67.
3. Greicius MD, Srivastava G, Reiss AL, Menon V. Default-mode network acti-
vity distinguishes Alzheimer’s disease from healthy aging: evidence from functio-
nal MRI. Proc Natl Acad Sci USA 2004;101:4637-42.
4. Kalpouzos G, Eustache F, de la Sayette V et al. Working memory and
FDG-PET dissociate early and late onset Alzheimer’s disease patients. J Neurol
2005;252:548-58.
5. Simons JS et Spiers HJ. Prefrontal and medial temporal lobe interactions in
long-term memory. Nat Rev Neurosci 2003;4:637-48.
6. McIntosh AR, Nyberg L, Bookstein FL, Tulving E. Differential functional
connectivity of prefrontal and medial temporal cortices during episodic memory
retrieval. Hum Brain Mapp 1997;5:323-7.
7. Ranganath C, Heller A, Cohen MX et al. Functional connectivity with the
hippocampus during successful memory formation. Hippocampus 2005;15:997-
1005.
8. Kilpatrick L et Cahill L. Amygdala modulation of parahippocampal and
frontal regions during emotionally influenced memory storage. Neuroimage
2003;20:2091-9.
9. Daselaar SM, Fleck MS, Dobbins IG et al. Effects of healthy aging on hip-
pocampal and rhinal memory functions: an event-related fMRI study. Cereb
Cortex 2006;16:1771-82.
10. Fellgiebel A, Müller MJ, Wille P et al. Color-coded diffusion-tensor-imaging
of posterior cingulate fiber tracts in mild cognitive impairment. Neurobiol Aging
2005;26:1193-8.
11. Kalus P, Slotboom J, Gallinat J et al. Examining the gateway to the lim-
bic system with diffusion tensor imaging: the perforant pathway in dementia.
Neuro-image 2006;30:713-20.
12. Kuroki N, Kubicki M, Nestor PG et al. Fornix integrity and hippocampal
volume in male schizophrenic patients. Biol Psychiatry 2006;60:22-31.
13. Persson J, Nyberg L, Lind J et al. Structure-function correlates of cognitive
decline in aging. Cereb Cortex 2006;16:907-15.
1
/
5
100%