cours 7

Scores d’alignement
Matrices de substitution
J.S. Bernardes/H. Richard

■ Score alignement = Σ scores événements élémentaires
(Match, Mismatch, Indel)
■ Amélioration du modèle!: pénalité linéaire des gaps (gap
open et gap extend)
■ Amélioration du modèle!: les matrices de substitution (=
Mismatch) => toutes les substitutions ne sont pas
équivalentes et donc pénalisées différemment
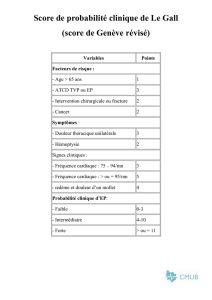
Score d'un alignement

Les matrices de substitution des acides
nucléiques
■ Alphabet à 4 lettres!: A,C,G,T
■ Matrice identité
• => pas d'amélioration du modèle,
• non adapté à l'évolution des séq nucléiques
■ Matrice transition/transvertion
• => pénalise davantage les
• transversions (purines <=> pyrimidines)
• que les
• transitions (pur <=> pur, pyr <=> pyr)
A C G T
A 1 0 0 0
C 0 1 0 0
G 0 0 1 0
T 0 0 0 1
A C G T
A 3 0 1 0
C 0 3 0 1
G 1 0 3 0
T 0 1 0 3

Les matrices de substitution des acides
aminés
■ Au cours de l’évolution:
❑ Des acides aminés sont remplacés «!préférentiellement!»
par d’autres
❑ Ils possèdent par exemple des propriétés physico-chimiques proches
❑ Des acides aminés sont plus conservés que d’autres
❑ Ils sont par exemple essentiels dans la structure 3D des protéines
(comme Tryptophane/W/Trp)

Les matrices de substitution des acides
aminés
■ Matrices basées sur les propriétés physico-chimique
des aa
o matrices d'hydrophobicité
o matrices des structures secondaires
o matrices basées sur comparaisons de protéines partagent la même structure 3-D
■ Matrices basées sur les substitutions entre aa au cours
de l'évolution
• Les «!log odds!»!: Sij = log [ qij / (pi.pj) ]
• qij = probabilité de la substitution i vers j
• pi = probabilité normalisée d’apparition du résidu i
• pj = probabilité normalisée d’apparition du résidu j
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
1
/
66
100%