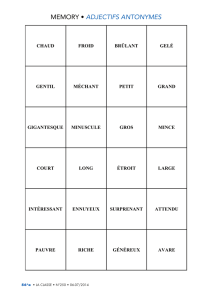
Présentée pour obtenir le grade de Docteur de l`Université de

THESE
Présentée pour obtenir le grade de
Docteur de l’Université de Strasbourg
Spécialité : Neurosciences
Par Joëlle Lopez
Dynamique mnésique du traitement de l’information spatiale
chez le Rat : approche systémique de la consolidation
4 Décembre 2009
Membres du jury
Jean-Christophe Cassel Directeur de thèse
Anne-Laurence Boutillier Rapporteur interne
Bruno Poucet Rapporteur externe
Pascal Roullet Rapporteur externe
Anne Pereira de Vasconcelos Membre invité
Bruno Bontempi Membre invité


Science is not meant to cure us of mystery, but to reinvent and reinvigorate it.
--- Robert Sapolsky, Why Zebras Don’t Get Ulcers, p. xii, 1994


Remerciements
En premier lieu, je souhaite remercier le Dr. Christian Kelche de m’avoir accueilli dans son
laboratoire où j’ai pu m’enrichir, tant sur le plan personnel que professionnel, ces quatre
dernières années. Merci également pour sa bonne humeur et nos discussions enjouées; ses
allers-retours dans le couloir en chantonnant ou en sifflotant ont su égayer plus d’une
journée…
Je tiens particulièrement à remercier mon directeur de thèse, le Pr. Jean-Christophe Cassel, de
m’avoir encadré tout au long de ce périple. Sa passion pour la Recherche et son énergie
débordante n’ont fait que renforcer ma motivation. Je le remercie d’avoir aiguisé mon sens
critique et de m’avoir poussé toujours plus loin dans mes réflexions. Merci également pour
son humour cynique et ses rires contagieux qui ont ponctué cette thèse de moments très
agréables !
Je remercie profondément le Dr. Anne Pereira de Vasconcelos, qui a co-encadré cette thèse.
Merci pour sa constante disponibilité, son sens de l’écoute et ses encouragements dans les
moments de frustration…Sans oublier les moments de joie partagés avec grand plaisir ! Nos
nombreuses discussions ont beaucoup contribué à mon épanouissement et à la construction de
cette thèse.
Un grand merci au Dr. Anne-Laurence Boutillier, au Dr. Bruno Poucet, au Dr. Pascal Roullet
et au Dr. Bruno Bontempi d’avoir accepté de faire partie de ce jury de thèse et d’avoir
consacré leur temps à la lecture de ce manuscrit.
Un grand merci également à Brigitte Cosquer et Karin Herbeaux-Geiger pour leurs doigts de
fées, leurs précieux conseils, et surtout pour leur bonne humeur et nos nombreux fous rires en
salle d’op et salle d’immuno !
Merci à Christophe, Hayat, Julie, Lucas, Marc-Antoine, Nour, PH et Sami pour tous ces
moments de complicité et nos nombreux petits délires. Sans eux cette aventure n’aurait pas
été aussi riche en couleurs…
Merci à Rodrigue pour son amitié et son soutien qui m’ont été essentiels tout au long de cette
thèse.
Merci à tous les membres et stagiaires du ex-LN2C/LINC Espla : Alexandra, Bernadette,
Catherine, Chantal, Christine, Daniel, Elise, Georges, Hélène, Marion, Michel, Monique,
Olivier, Patrick, Philippe, Sarah S., Sarah P, Vincent et Virginie. Ils ont su construire la bonne
ambiance qui règne au laboratoire, et ont ainsi été une sorte de seconde famille pour moi
pendant ces quatre années passées en leur compagnie. Pour cela je leur en serai toujours
reconnaissante.
Merci à Jenni pour son amitié sans limites et tous les moments complices qu’on a pu partagé.
Merci aussi à tous les Doctoneuro et à la grande solidarité qui existe au sein de ce groupe. Je
n’oublierai jamais ce voyage en Estonie, si riche en émotions !
Enfin, merci à ma famille pour tout leur amour et leur soutien indéfectibles.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
1
/
220
100%