Cécile, Madeleine DEVRED - Thèses

ÉCOLE NATIONALE VETERINAIRE D’ALFORT
Année 2004
ADAPTATIONS ANATOMO-PHYSIOLOGIQUES ET
COMPORTEMENTALES A LA REPRODUCTION EN
ANTARCTIQUE CHEZ LE MANCHOT EMPEREUR
THESE
Pour le
DOCTORAT VETERINAIRE
Présentée et soutenue publiquement devant
LA FACULTE DE MEDECINE DE CRETEIL
le……………
par
Cécile, Madeleine DEVRED
Né (e) le 21 avril 1977 à Clamart (Hauts-de-Seine)
JURY
Président : M.
Professeur à la Faculté de Médecine de CRETEIL
Membres
Directeur : Mme Hélène COMBRISSON
Professeur Physiologie-Thérapeutique à l’ENVA
Assesseur : M. Bertrand DEPUTTE
Professeur d’Ethologie à l’ENVA

LISTE DES MEMBRES DU CORPS ENSEIGNANT
Directeur : M. le Professeur COTARD Jean-Pierre
Directeurs honoraires : MM. les Professeurs PARODI André-Laurent, PILET Charles
Professeurs honoraires: MM. BORDET Roger,BUSSIERAS Jean,LE BARS Henri, MILHAUD Guy,ROZIER Jacques,THERET Marcel,VUILLAUME Robert
DEPARTEMENT DES SCIENCES BIOLOGIQUES ET PHARMACEUTIQUES (DSBP)
Chef du département : M. BOULOUIS Henri-Jean, Professeur - Adjoint : M. DEGUEURCE Christophe, Professeur
-UNITE D’ANATOMIE DES ANIMAUX DOMESTIQUES
Mme CREVIER-DENOIX Nathalie, Professeur*
M. DEGUEURCE Christophe, Professeur
Mlle ROBERT Céline, Maître de conférences
M. CHATEAU Henri, AERC
-UNITE DE PATHOLOGIE GENERALE , MICROBIOLOGIE,
IMMUNOLOGIE
Mme QUINTIN-COLONNA Françoise, Professeur*
M. BOULOUIS Henri-Jean, Professeur
Mme VIALE Anne-Claire, Maître de conférences
-UNITE DE PHYSIOLOGIE ET THERAPEUTIQUE
M. BRUGERE Henri, Professeur *
Mme COMBRISSON Hélène, Professeur
M. TIRET Laurent, Maître de conférences
-UNITE DE PHARMACIE ET TOXICOLOGIE
Mme ENRIQUEZ Brigitte, Professeur *
Mme HUYNH-DELERME, Maître de conférences contractuel
M. TISSIER Renaud, Maître de conférences
- UNITE D’HISTOLOGIE , ANATOMIE PATHOLOGIQUE
M. CRESPEAU François, Professeur *
M. FONTAINE Jean-Jacques, Professeur
Mme BERNEX Florence, Maître de conférences
Mme CORDONNIER-LEFORT Nathalie, Maître de conférences
-UNITE DE BIOCHIMIE
M. BELLIER Sylvain, Maître de conférences*
M. MICHAUX Jean-Michel, Maître de conférences
- UNITE DE VIROLOGIE
M. ELOIT Marc, Professeur *
Mme ALCON Sophie, Maître de conférences contractuel
-DISCIPLINE : PHYSIQUE ET CHIMIE BIOLOGIQUES ET
MEDICALES
M. MOUTHON Gilbert, Professeur
-DISCIPLINE : BIOLOGIE MOLECULAIRE
Melle ABITBOL Marie, Maître de conférences contractuel
-DISCIPLINE : ETHOLOGIE
M. DEPUTTE Bertrand, Professeur
DEPARTEMENT D’ELEVAGE ET DE PATHOLOGIE DES EQUIDES ET DES CARNIVORES (DEPEC)
Chef du département : M. FAYOLLE Pascal, Professeur - Adjointe : Mme BEGON Dominique , Professeur
-UNITE DE MEDECINE
M. POUCHELON Jean-Louis, Professeur*
M. CLERC Bernard, Professeur
Mme CHETBOUL Valérie, Professeur
M. MORAILLON Robert, Professeur
M. BLOT Stéphane, Maître de conférences
M. ROSENBERG Charles, Maître de conférences contractuel
Melle MAUREY Christelle, Maître de conférences contractuel
- UNITE DE CLINIQUE EQUINE
M. DENOIX Jean-Marie, Professeur *
M. TNIBAR Mohamed, Maître de conférences contractuel
M. AUDIGIE Fabrice, Maître de conférences
Mme DESJARDINS-PESSON Isabelle, Maître de confér..contractuel
-UNITE DE REPRODUCTION ANIMALE
M. MIALOT Jean-Paul, Professeur * (rattaché au DPASP)
M. NUDELMANN Nicolas, Maître de conférences
Mme CHASTANT-MAILLARD Sylvie, Maître de conférences
(rattachée au DPASP )
M. FONTBONNE Alain, Maître de conférences
M. REMY Dominique, Maître de conférences (rattaché au DPASP)
Melle CONSTANT Fabienne, AERC (rattachée au DPASP)
- UNITE DE PATHOLOGIE CHIRURGICALE
M. FAYOLLE Pascal, Professeur *
M. MAILHAC Jean-Marie, Maître de conférences
M. MOISSONNIER Pierre, Professeur
Mme VIATEAU-DUVAL Véronique, Maître de conférences
M. DESBOIS Christophe, Maître de conférences
Mlle RAVARY Bérangère, AERC (rattachée au DPASP)
M. ZILBERSTEIN Luca, Maître de Conférences contractuel
M. HIDALGO Antoine, Maître de Conférences contractuel
- UNITE DE RADIOLOGIE
Mme BEGON Dominique, Professeur*
M. RUEL Yannick, AERC
- UNITE DE PARASITOLOGIE ET MALADIES PARASITAIRES
M. CHERMETTE René, Professeur *
M. POLACK Bruno, Maître de conférences
M. GUILLOT Jacques, Professeur
Melle MARIGNAC Geneviève, Maître de conférences contractuel
M. PARAGON Bernard, Professeur (rattaché au DEPEC)
M. GRANDJEAN Dominique, Professeur (rattaché au DEPEC)
DEPARTEMENT DES PRODUCTIONS ANIMALES ET DE LA SANTE PUBLIQUE (DPASP)
Chef du département : M. CERF Olivier, Professeur - Adjoint : M. BOSSE Philippe, Professeur
-UNITE DES MALADIES CONTAGIEUSES
M. TOMA Bernard, Professeur
M. BENET Jean-Jacques, Professeur*
Mme HADDAD H0ANG XUAN Nadia, Maître de confér.contractuel
M. SANAA Moez, Maître de conférences
-UNITE D’HYGIENE ET INDUSTRIE DES ALIMENTS
D’ORIGINE ANIMALE
M. BOLNOT François, Maître de conférences *
M. CARLIER Vincent, Professeur
M. CERF Olivier, Professeur
Mme COLMIN Catherine, Maître de conférences
M. AUGUSTIN Jean-Christophe, Maître de conférences
- UNITE DE ZOOTECHNIE, ECONOMIE RURALE
M. BOSSE Philippe, Professeur
M. COURREAU Jean-François, Professeur*
Mme GRIMARD-BALLIF Bénédicte, Maître de conférences
Mme LEROY Isabelle, Maître de conférences
M. ARNE Pascal, Maître de conférences
M. PONTER Andrew, Maître de conférences
- UNITE DE PATHOLOGIE MEDICALE DU BETAIL ET DES
ANIMAUX DE BASSE-COUR
Mme BRUGERE-PICOUX Jeanne, Professeur
M.MAILLARD Renaud, Maître de conférences associé
M. MILLEMANN Yves, Maître de conférences*
M. ADJOU Karim, Maître de conférences
Ingénieurs Professeurs agrégés certifiés (IPAC) : * Responsable de l’Unité
Mme CONAN Muriel, Professeur d’Anglais
Mme CALAGUE, Professeur d’Education Physique AERC : Assistant d’Enseignement et de Recherche Contractuel

J’adresse tous mes remerciements
A Monsieur le Professeur de la faculté de Créteil,
qui me fait l’honneur de présider le jury de cette thèse.
Au Professeur Hélène Combrisson,
pour l’attention qu’elle à apportée à l’examen de mon travail et
qui me fait l’honneur d’être la directrice de cette thèse.
Au Professeur Bertrand Deputte,
pour l’intérêt qu’il a porté à mon travail et
qui me fait l’honneur d’être l’assesseur de cette thèse.
A mes proches, famille et amis,
pour leur soutien, leur confiance et leur patience dans tout ce que j’ai entrepris.

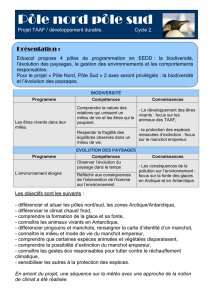
ADAPTATIONS ANATOMO-PHYSIOLOGIQUES ET
COMPORTEMENTALES A LA REPRODUCTION EN ANTARCTIQUE
CHEZ LE MANCHOT EMPEREUR
NOM et Prénom : DEVRED Cécile
Résumé : Lors de son cycle reproducteur, le manchot empereur subit des températures comprises entre 0 et -50°C,
avec des vents souvent supérieurs à 200 Km/h. Ce travail a pour objectif d’illustrer les adaptations de cette espèce à
son milieu au moyen de photos et d’enregistrements audio-visuels sur un support cédérom.
Le manchot empereur maintient son homéothermie grâce à une faible surface corporelle relative et un plumage dont
la structure assure plus de 85 % de l’isolation thermique. Dans les parties du corps les moins isolées thermiquement
(ailerons et pattes), les systèmes vasculaires d’échangeurs à contre-courant permettent une économie d’énergie par
réduction des déperditions thermiques. L’augmentation des phases de sommeil lent et la faible activité du manchot
empereur sont également des moyens de diminuer sa dépense énergétique. Ces adaptations anatomo-physiologiques
sont complétées par un comportement thermorégulateur social des couveurs qui transportent leur œuf sur leurs
pattes. Le regroupement des oiseaux en formations serrées leur permet de maintenir un métabolisme moyen
inférieur de 25 % à celui mesuré chez des oiseaux isolés à neutralité thermique. Seul oiseau de mer à ne pas
posséder d’emplacement de reproduction fixe, la reconnaissance individuelle entre partenaires ou entre parent et
poussin est assurée par le chant. Le maintien des liens du couple est assuré par le silence préposital et la parade
mutuelle. La courbette permet la synchronisation des partenaires, indispensable lors de l’échange de l’œuf et du
poussin sous ce climat.
Mots clés :
Jury :
Président : Pr.
Directeur : Pr. Hélène COMBRISSON
Assesseur : Pr. Bertrand DEPUTTE
Adresse de l’auteur : Cécile DEVRED
37, rue des Clos Beauregards
92 500 RUEIL MALMAISON

ANATOMO-PHYSIOLOGICAL AND BEHAVIORAL ADAPTATIONS
TO ANTARCTIC BREEDING IN EMPEROR PENGUIN
SURNAME : DEVRED
Given name : Cécile
Summary : During his breeding cycle, the emperor penguin has to deal with ambiant temperatures between 0 and -
50°C, and wind velocities often higher than 200 Km/h. The aim of this work is to illustrate the adaptations of this
species to its environment by the mean of photographies and audio-visual records on a cederom aid.
The emperor penguin maintains his homeothermy thanks to a relatively low body surface area and to a plumage
structure which provides more than 85 % of its thermal insulation. In the less insulated parts (wings and feet),
vascular counter-current heat exchangers enable energy saving by reducing themal losses. The increase of slow
wave sleep and the low activity level also contribute to decrease the energetic expends of the emperor penguin.
Those anatomo-physiological adaptations are completed by the social thermoregulatory behaviour of breeder which
carry its egg on its feets. By packing in tiny groups, the birds maintain a metabolic rate 25 % below that measured in
isolated birds at thermoneutrality. In this non-nesting seabird species, individual recognition between partners or
parents and chick is assured by the call. Preposital silence and courtship display help to maintain pair bondings. The
mutual display bowing unable the synchronization between partners, essential during the egg or chick exchange
under this climate.
Keywords :
Jury :
Président : Pr.
Director : Pr. Hélène COMBRISSON
Assesor : Pr. Bertrand DEPUTTE
Author’s address: Cécile DEVRED
37, rue des Clos Beauregards
92 500 RUEIL MALMAISON
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
1
/
59
100%