Télécharger le fichier

1
FAIRE LE POINT
Messages et messagers dans les corrélations hormonales et nerveuses
Le fonctionnement coordonné de l’organisme repose sur l’existence de deux modalités de
communication, la communication nerveuse et la communication hormonale ; ces
deux modes de communication sont gouvernés par le même principe général : émission
d’un signal, transport, réception, transduction, intégration, fin du signal.
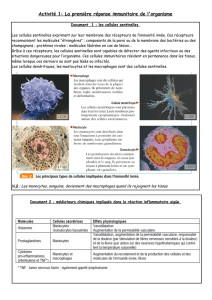
La communication hormonale repose sur la libération de molécules, les hormones, dans
le milieu extracellulaire. Seules les cellules qui possèdent des récepteurs spécifiques à
ces molécules réagissent à leurs présences. On appelle hormone une substance produite
par des cellules, libérée en faible quantité dans le sang et qui agit à distance sur certaines
catégories de cellules, les cellules cibles. Selon la distance parcourue par ces molécules,
on distingue les communications autocrines, paracrines (facteurs paracrines) et
endocrines (hormones). La communication neurohormonale correspond à la sécrétion
par un neurone d’une molécule (neurohormone) dans le sang.
Des hormones de nature différente, en se fixant sur des récepteurs différents, produisent
les mêmes effets. Ainsi, le glucagon sécrété par le pancréas et l’adrénaline libérée par
les glandes médullosurrénales conduisent à la libération de glucose par le foie.
Les cellules cibles des hormones se caractérisent par la présence de récepteurs
membranaires, hyaloplasmiques ou nucléaires qui fixent l’hormone avec une grande
affinité. Le changement de conformation du récepteur hormonal est responsable des
effets de l’hormone. La réponse de la cellule cible dépend de la concentration
hormonale, du nombre de récepteurs exprimés par la cellule, de leur affinité pour
l’hormone et de la concentration des protéines cibles dans la cellule.
Le système nerveux se construit progressivement au cours du développement par la
croissance et les contacts qu’établissent ses cellules spécialisées, les neurones. Il relie les
différents organes aux centres nerveux. Un centre nerveux est une régionalisation qui
comprend des synapses, c’est-à-dire des zones de contact entre différents neurones. La
communication par le système nerveux repose sur un câblage préétabli. Le système
nerveux comprend le système nerveux central qui reçoit des voies sensorielles et donne
naissance à des voies motrices, il est responsable de l’activité musculaire squelettique
volontaire et involontaire (réflexe d’évitement) et le système nerveux autonome
(système nerveux sympathique, système nerveux parasympathique) qui contrôle
l’activité des viscères et du système cardiovasculaire.
Le long de l’axone, le message nerveux est codé en modulation de la fréquence des
potentiels d’action. Arrivé à une synapse, il est codé en concentration de
neurotransmetteurs qui ouvrent des canaux ioniques qui assurent éventuellement la
transmission du message au neurone suivant. Les neurones du système nerveux
sympathique libèrent à leurs extrémités de la noradrénaline et un peu l’adrénaline qui
agissent sur des cellules cibles au même titre que ces deux molécules quand elles sont
libérées dans le sang (hormone) par les glandes médullosurrénales. Les neurones du
système nerveux parasympathique libèrent à leurs extrémités de l’acétylcholine dont
les effets sont souvent antagonistes de ceux qui sont provoqués par l’adrénaline et la
noradrénaline.
On appelle neurotransmetteur une molécule qui est libérée par l’extrémité d’un neurone
à proximité d’une autre cellule (neurone, cellule musculaire) et qui se fixe sur des
récepteurs spécifiques. L’ensemble formé par l’espace entre les deux surfaces cellulaires
et l’espace qui les sépare est une synapse. Selon le type de cellules et la nature des

2
récepteurs, un même neurotransmetteur provoque des effets différents, tel est l’exemple
de l’acétylcholine qui, libérée par le système nerveux parasympathique, ralentit le
cœur ou libéré par des neurones moteurs, entraîne la contraction des cellules musculaires
striées.
L’action de ces deux systèmes de communication est complémentaire, le système nerveux
a un effet rapide mais de courte durée alors que la communication hormonale demande
plus de temps pour s’établir mais les effets durent plus longtemps.

3
FAIRE LE POINT
Mode d’action cellulaire des neurotransmetteurs et des hormones
Les propriétés physicochimiques des neurotransmetteurs et des hormones conditionnent
leurs modalités d’action. Il est ainsi commode de distinguer les molécules hydrophiles
qui ne traversent pas les membranes plasmiques et les molécules lipophiles qui les
traversent. Ces propriétés de solubilité conditionnent la localisation de leurs récepteurs.
On distingue ainsi les hormones et les neurotransmetteurs qui se fixent sur des récepteurs
transmembranaires de ceux qui se lient à des récepteurs intracellulaires.
L’adrénaline, la noradrénaline et le glucagon se lient à des récepteurs couplés à un
système de transduction du message appelé protéine G. Il existe 4 catégories de
récepteurs fixant l’adrénaline et la noradrénaline : les récepteurs α1, α2, β1 et β2. La
répartition de ces récepteurs dépend des cellules. La noradrénaline se fixe
préférentiellement sur les récepteurs α alors que l’adrénaline présente une plus grande
affinité pour les récepteurs β.
Les récepteurs α1 dominent sur les cellules musculaires lisses des vaisseaux sanguins
autres que ceux du cœur et des artérioles des muscles ; les récepteurs β1 sont représentés
sur les myocytes striés cardiaques et sur les cellules musculaires lisses des vaisseaux
coronaires et les récepteurs β2 sur les cellules musculaires lisses des autres artérioles et
sur la membrane des hépatocytes et des myocytes squelettiques.
La fixation de ces molécules sur les récepteurs β des cellules musculaires lisses aboutit à
l’activation d’une protéine membranaire, la protéine G dont une sous-unité mobile se
déplace latéralement dans le feuillet cyctosolique et active soit l’adénylate cyclase
(récepteurs β1 et β2), soit l’inhibe (récepteur α2), soit la phospholipase C (récepteur
α1). L’activation de l’adénylate cyclase conduit à une augmentation du taux d’AMPc qui
se fixe sur une protéine kinase (PKA) qui était inactive. Rendue active, la PKA
phosphoryle des protéines cibles qui dépendent des cellules cibles. Les effets de
l’adrénaline et de la noradrénaline sur les récepteurs β correspondent aux protéines
cellulaires phosphorylées par la PKA. C’est la voie de transduction de l’adénylate
cyclase qui conduit à la formation d’un second messager l’AMPc.
L’activation de la phospholipase C (récepteurs α1) conduit à l’hydrolyse d’un lipide du
feuillet cytosolique, le phosphatidyl-inositol 4,5-bisphosphate (PIP2) en 1,2
diacylglycérol (DAG) qui reste dans le feuillet et en inositol 1,4,5-trisphosphate (IP3)
soluble qui gagne le hyaloplasme. L’IP3 se fixe sur des canaux Ca2+ situés sur la
membrane du réticulum et les ouvre, le Ca
2+ gagne le hyaloplasme, il se fixe sur une
protéine kinase (PKC), l’ensemble se lie au DAG qui l’active. L’IP3, le DAG et le Ca2+
sont des seconds messagers. La PKC devenue active phosphoryle ses protéines cibles.
Les effets de l’hormone correspondent aux activités de protéines ainsi phosphorylées. Par
ailleurs, le Ca2+ se fixe à une autre protéine, la calmoduline. La calmoduline présente de
très nombreuses protéines cibles parmi lesquelles on note les CAM-kinases et les
phosphodiestérases dont l’activation diminue la quantité d’AMPc et arrête la réponse.
Ces systèmes de transduction conduisent au phénomène d’amplification de la réponse
hormonale. Tant que la protéine G est liée à l’adénylate cyclase ou à la phospholipase C,
ces molécules produisent des seconds messagers qui activent la PKA et la PKC. Chacune
de ces molécules phosphoryle de nombreux substrats. Ainsi, quelques molécules
d’hormones se fixant sur quelques récepteurs suffisent à donner une réponse de grande
ampleur.

4
L’acétylcholine est un neurotransmetteur qui est soit libéré par le système nerveux
parasympathique, soit par le système nerveux périphérique. L’acétylcholine libérée
par les fibres parasympathiques se fixe sur les récepteurs muscariniques des myocytes
cardiaques et, par l’intermédiaire d’une protéine G, entraîne l’ouverture de canaux K+,
la membrane s’hyperpolarise ce qui ralentit le rythme cardiaque. L’acétylcholine libérée
par les terminaisons des motoneurones se lient aux récepteurs nicotiniques qui s’ouvrent
directement et laisse entrer du Na+ qui dépolarise la membrane et déclenche
éventuellement la naissance d’un potentiel d’action musculaire.
Les hormones liposolubles (thyroxine et hormones dérivées du cholestérol : cortisol,
testostérone, œstrogènes) franchissent les membranes de toutes les cellules et, dans les
cellules cibles, augmentent la synthèse protéique. Ces hormones stéroïdes se fixent sur
des récepteurs cytosoliques qui se dimérisent dans le noyau. Ces homodimères se lient
à des séquences d’ADN de nature palindromique appelées éléments de réponses à
l’hormone. La fixation de ce dimère augmente la stabilité du complexe de transcription
et donc le taux de transcription des gènes.
Les récepteurs à la thyroxine, certains sont sous forme d’homodimères (TR-TR) alors
que d’autres sont sous forme d’hétérodimères (TR-RXR), sont liés aux séquences
palindromiques des gènes qu’elle régule. En l’absence de thyroxine, les hétérodimères
recrutent des molécules (corépresseurs) qui maintiennent la chromatine à l’état
condensé. En présence de thyroxine, l’hétérodimère change de forme, de nouvelles
molécules se fixent et assurent le déroulement de la chromatine.
Pour les deux types d’hormones, l’importance de la réponse dépend de la concentration
en hormones et du nombre de récepteurs. Le nombre de récepteurs est contrôlé par le
taux de transcription des gènes qui les codent.
Les modalités d’arrêt de la réponse à la fixation d’un ligand dépendent des cellules.
Pour les hormones hydrophiles, la diminution de l’affinité de la protéine G pour le
récepteur, la séparation de la protéine G de l’enzyme membranaire, l’action de la
phosphodiestérase mettent fin à la réponse hormonale. Pour les neurotransmetteurs,
l’inactivation par des enzymes spécifiques (MAO pour la noradrénaline,
acétylcholinestérase pour l’acétylcholine), la recapture par la terminaison axonique , la
diffusion dans la fente mettent fin à la réponse.
Sur le plus long terme, la diminution de l’affinité du récepteur couplé à une protéine G à
la suite de sa phosphorylation, la fixation d’arrestine sur le récepteur ainsi phosphorylé
et son endocytose mettent fin à la réponse.
Pour les hormones lipophiles, la durée d’action est variable. Chez les Amphibiens,
l’action de la thyroxine sur le génome dure toute la vie ; en revanche, la fixation des
récepteurs aux glucocorticoïdes est temporaire et les gènes ainsi activés ne sont transcrits
que peu de temps.

5
FAIRE LE POINT
Genèse et propagation des messages nerveux
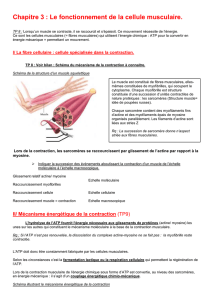
Le circuit nerveux le plus simple est celui du réflexe myotatique qui contrôle l’état de
contraction du muscle. Tout muscle est soumis à son propre poids qui l’étire. Cet
étirement est détecté par des structures sensorielles (fuseaux neuromusculaires) qui
génèrent un message nerveux sensoriel qui voyage dans un neurone passant par la racine
dorsale de la moelle épinière. Ce neurone établit une synapse avec un interneurone relié
à un motoneurone innervant le muscle antagoniste et directement à un motoneurone du
muscle étiré. Ces deux neurones sortent par la racine ventrale. Il s’ensuit une légère
contraction du muscle étiré qui maintient un certain tonus musculaire. Les étapes du
réflexe myotatique sont : naissance et codage de l’intensité du stimulus, propagation,
transmission, intégration, genèse d’un nouveau message, propagation, transmission à
la cellule effectrice.
Il existe une différence de potentiel électrique de part et d’autre de la membrane
plasmique de toute cellule vivante. Cette différence est due à une perméabilité sélective
de la membrane et à l’existence de transports actifs. Les ions les plus concentrés dans le
milieu extracellulaire sont le Na+, le Ca2+ et le Cl−, en revanche, le K+ est plus concentré
dans le milieu intracellulaire. La cellule laisse sortir plus de K+ qu’elle ne laisse entrer de
cations, son côté cytosolique est donc négatif par rapport à sa surface. Le déplacement
des ions de part et d’autre de la membrane est contrôlé par la force osmotique et la force
électrique mais aussi par la perméabilité membranaire. Au repos, le flux net de Na+ est
entrant, le flux net de K+ est sortant et le flux net de Cl
− est nul. La pompe Na+/K+
ATPase éjecte 3 Na+ et prélève 2 K+ qui compense les 3 Na+ entrés et les 2 K+ sortis dans
le sens de leurs gradients électrochimiques respectifs.
Les neurones sont des cellules excitables, ils possèdent une catégorie spécifique de
canaux, les canaux voltage-dépendants. Ils propagent une onde de dépolarisation sans
que celle-ci ne s’atténue. Cette onde est le potentiel d’action. Il se traduit par une
dépolarisation, une repolarisation et une hyperpolarisation d’une amplitude d’environ
100 mV et d’une durée de quelques millisecondes. Cette onde est due à un changement de
conductance ionique. La dépolarisation est provoquée par une augmentation transitoire
de la perméabilité au Na+, la repolarisation et l’hyperpolarisation à une élévation de la
conductance au K+.
L’ouverture séquentielle des canaux voltage-dépendants est responsable de ces
changements de conductance. Ces canaux s’ouvrent en fonction de la valeur du potentiel
de membrane de la membrane qui les entoure. Le potentiel de repos est de l’ordre de − 70
mV ; quand il atteint environ − 40 mV, les canaux Na+ voltage-dépendants s’ouvrent
massivement et le Na+ entre dans la cellule sous la force de son gradient
électrochimique, le potentiel atteint alors 35 mV, les canaux K+ voltage-dépendants
s’ouvrent, le K+ sort, il repolarise et hyperpolarise la membrane.
La technique du patch-clamp associée à la connaissance de la structure tridimensionnelle
des canaux voltage-dépendants a permis de comprendre leur fonctionnement et leur
contribution à la formation d’un potentiel d’action. L’ouverture et la fermeture de ses
canaux sont contrôlées par des hélices transmembranaires tensiosensibles chargées
positivement qui se déplacent en fonction de la valeur de la polarisation membranaire.
Dans un axone non stimulé, la plupart des canaux voltage-dépendants sont fermées, mais
s’ouvrent si la membrane se dépolarise au-delà d’une valeur seuil. Les canaux voltage-
dépendants possèdent une période réfractaire qui les empêche de s’ouvrir
immédiatement après s’être ouverts. Cette période réfractaire est responsable de la
conduction unidirectionnelle du potentiel d’action dans les conditions physiologiques.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%