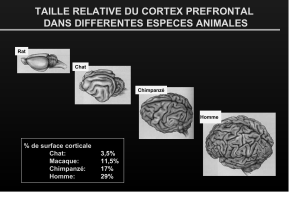
Syndrome dysexécutif cognitif

doi: 10.1684/nrp.2009.0005
Syndrome dysexécutif cognitif :
un déficit de l’administrateur
de la mémoire de travail ?
The cognitive dysexecutive
syndrome: an impairment
of the central executive
of working memory?
Résumé Le syndrome dysexécutif cognitif est secondaire à l’alté-
ration des processus cognitifs permettant d’élaborer et de
contrôler volontairement les comportements dirigés vers un but. Ces processus cognitifs
regroupent de nombreuses macrofonctions (planification, raisonnement, découverte de
règles, etc.), reposant elles-mêmes sur des opérations élémentaires appartenant à la mémoire
de travail. La conception psychologique de la mémoire de travail isole les systèmes de
maintien de l’information en mémoire à court terme (les systèmes tampons) du système de
gestion exécutive (l’administrateur central), manipulant et utilisant l’information maintenue en
représentation mentale. Ce modèle psychologique a une pertinence anatomo-fonctionnelle,
car il permet de dissocier les fonctions des cortex associatifs rétrorolandiques, indispensables
pour le maintien en mémoire à court terme, du cortex préfrontal latéral dont les fonctions se
superposent grandement à l’administrateur central de la mémoire de travail. Toutefois, si le
syndrome dysexécutif cognitif correspond le plus souvent à une altération de l’administrateur
central (terme qui tend progressivement à être remplacé par celui de « contrôle cognitif »), lui-
même secondaire à un dysfonctionnement préfrontal, une vision connexionniste et interactive
des fonctions cérébrales suppose que l’intégrité des fonctions exécutives dépend d’un réseau
plus vaste, incluant les cortex associatifs rétrorolandiques, le cortex frontal orbitaire et médian
et les ganglions de la base.
Mots clés : cortex préfrontal
•
contrôle cognitif
•
fonctions exécutives
•
IRMf
Abstract The cognitive dysexecutive syndrome is secondary to
the impairment of the cognitive processes required to
voluntary elaborate and control goal-directed behaviors. Planning, reasoning, rule fin-
ding, abstraction are some of the cognitive processes gathered under the concept of exe-
cutive functions. They rely on more elementary operations that typically belong to wor-
king memory. According to the classical Baddeley’s psychological model, working
memory is composed of buffer systems that maintain information in short-term memory
and of a central executive module, essential to manipulate mental representations to pre-
pare the forthcoming voluntary action. This psychological model is of interest for clini-
cians and neuroscientists because it can be related to a relevant anatomical-functional
dissociation between retro-rolandic association cortices and the dorsolateral prefrontal
cortex. Indeed, a large set of data, mostly from the monkeys and from imaging studies in
human, supports the idea that the retro-rolandic association areas are involved in the
short-term maintenance of information. In these posterior cortical brain regions, mainte-
nance is topographically organized according to the domain of the material maintained
Article de synthèse
Rev Neuropsychol
2009 ; 1 (1) : 34-41
Correspondance :
R. Lévy
Richard Lévy
Service de neurologie,
Hôpital Saint-Antoine, Paris ;
et Inserm U610,
Hôpital de la Pitié-Salpêtrière, Paris
R
EVUE DE NEUROPSYCHOLOGIE
N
EUROSCIENCES COGNITIVES ET CLINIQUES
34
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

in working memory (visuospatial, objects features, verbal…). The dorsolateral prefrontal
cortex is mostly involved in the central executive (or according to a more recent naming,
the “cognitive control”). How the dorsolateral prefrontal cortex is organized to support
its functions is still a matter of debate. Eventually, it is important to keep in mind, in a
modern and connexionnist conception of brain functions, that a vast neural network
(including retro-rolandic association cortices, orbital-medial prefrontal cortex and the
basal ganglia) is essential to support executive functions and cognitive control.
Key words: prefrontal cortex
•
executive functions
•
cognitive control
•
functional imaging studies
L’adaptation de l’homme à son environnement
nécessite l’apprentissage de règles physiques et
sociales bien établies (« si je m’approche du feu,
je me brûle. J’évite donc de m’en approcher »…). Ces règles
de base sont nécessaires pour la survie, le maintien de
l’homéostasie et la régulation minimale des comportements
sociaux. Ces règles rigides reposent sur l’apprentissage
associatif entre une connaissance ou un comportement et
son résultat invariant. Ces connaissances, certes indispen-
sables, demeurent insuffisantes pour l’adaptation pleine de
l’individu au monde complexe et mouvant dans lequel
nous évoluons. L’adaptation à des situations nouvelles ou
suffisamment complexes nous oblige à établir des compor-
tements régis par la mise en jeu de stratégies inédites et
planifiées, se dégageant des comportements automatiques,
réflexifs et routiniers. La flexibilité mentale, la résistance
aux interférences, le partage des ressources attentionnelles,
la découverte et l’apprentissage de nouvelles règles, la pla-
nification, le raisonnement, les stratégies pour réactiver les
traces mnésiques, la conceptualisation et l’abstraction sont
quelques-unes des fonctions mentales engagées dans les
processus adaptatifs face à un monde en mouvement per-
manent et aux règles complexes. Elles appartiennent classi-
quement aux fonctions exécutives. Dans un sens restrictif,
les fonctions exécutives sont l’ensemble des opérations
mentales requises pour la réalisation d’une tâche cognitive
complexe. Selon Miyake et al. [1], elles sont organisées
autour de trois fonctions mentales plus élémentaires : la
manipulation et l’actualisation des représentations menta-
les, le renversement de règles préétablies et l’inhibition
des réponses réflexives. Dans un sens plus large, les fonc-
tions exécutives peuvent être définies par l’ensemble des
fonctions mentales impliquées dans l’élaboration et le
contrôle de comportements volontaires, dirigés vers un
but [2]. En d’autres termes, toutes les opérations mentales
concourant à la réalisation volontaire de l’action font partie
des fonctions exécutives. Il est donc possible d’y inclure
toutes les fonctions mentales permettant de créer le but et
d’impulser le besoin d’agir (il s’agit des fonctions de volition
incluant les traitements affectifs poussant à agir comme la
prise en compte des renforcements positifs et négatifs
potentiels), toutes les fonctions nécessaires à élaborer le
plan d’action, une fois le but choisi (il s’agit des étapes
cognitives de construction de l’action), des étapes de
préparation motrice et d’exécution de l’action volontaire
et des processus nécessaires de rétrocontrôles cognitif et
affectif permettant de vérifier si le comportement en cours
est valide ou s’il doit être modifié [3]. Ainsi, le syndrome
dysexécutif peut être d’ordre cognitif ou socio-affectif.
Dans cet article, nous nous limiterons à discuter des bases
neuropsychologiques et neurales du versant cognitif du
syndrome dysexécutif, c’est-à-dire principalement des opé-
rations mentales permettant de construire un plan d’action
dirigé vers un but (nous mettrons de côté le versant affectif
ou social des fonctions exécutives).
La constitution d’un plan d’action repose sur des opérations
élémentaires telles que l’élaboration et le maintien d’un but
et de ses étapes intermédiaires, la programmation de la
séquence d’actions à effectuer et l’inhibition des actions
inappropriées. Certaines de ces opérations élémentaires
peuvent être regroupées sous le concept de mémoire de
travail. Le concept de mémoire de travail prend sa source
dans le champ de la psychologie cognitive sous l’impulsion
des travaux d’Atkinson et Shiffrin [4], puis de Baddeley [5].
Une définition de la mémoire de travail, généralement
admise par ceux utilisant l’approche expérimentale chez
le singe et l’imagerie fonctionnelle chez l’homme, est la
suivante : il s’agit d’un ensemble d’opérations mentales per-
mettant le maintien, la manipulation et l’utilisation de
représentations internes d’informations provenant du
monde extérieur ou du stock mnésique en prévision d’une
action [6]. Dans la définition proposée ci-dessus, la
mémoire de travail regroupe au moins deux ensembles de
processus cognitifs distincts mais interdépendants : la capa-
cité de maintenir à court terme une représentation, et la
gestion exécutive, c’est-à-dire l’utilisation mentale en vue
de l’action, des informations maintenues en représentation
interne. Cette fragmentation de la mémoire de travail
recoupe en grande partie le modèle psychologique de Bad-
deley [5] postulant à l’existence, d’une part, de systèmes
« tampons » permettant le stockage sous une forme
« active » de l’information et, d’autre part, d’un « adminis-
trateur central », gestionnaire allouant les ressources atten-
tionnelles et manipulant l’information maintenue active-
ment en mémoire à court terme. Dans sa finalité, la
mémoire de travail est très différente de la mémoire décla-
rative à long terme, car sa fonction n’est pas dirigée vers le
stockage (même s’il s’agit d’une étape obligatoire) mais vers
la réalisation d’actions à court terme. Comme le souligne, le
Article de synthèse
R
EVUE DE NEUROPSYCHOLOGIE
N
EUROSCIENCES COGNITIVES ET CLINIQUES
35
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

psychologue Baddeley [5], elle correspond à une interface
entre la mémoire et l’action.
L’administrateur central de la mémoire de travail repré-
sente un ensemble de processus actif sur les représentations
mentales maintenues en mémoire à court terme. Théorique-
ment, si ce module de la mémoire de travail était altéré, il
pourrait entraîner un syndrome dysexécutif d’intensité
sévère mais probablement dissocié. En effet, son altération
affecterait principalement l’ensemble des fonctions néces-
sitant la manipulation et l’actualisation des représentations
mentales en mémoire de travail. Il s’agit de la planification,
du raisonnement déductif, du calcul mental, de l’apprentis-
sage de procédures motrices ou cognitives séquentielles, de
l’élaboration de règles basées sur plusieurs paramètres, de
la réalisation concomitante de deux tâches cognitives, de la
navigation spatiale dans un espace représenté mentale-
ment, de la lecture de textes complexes ou narratifs, etc.
Toutefois, d’autres aspects des fonctions exécutives reste-
raient relativement préservés : il s’agit des capacités de
maintien des représentations mentales ne nécessitant pas
de réorganisation de l’information (capacités reposant sur
les systèmes « tampon »), les renversements de règles, les
capacités d’inhibition des actions réflexives, archaïques
ou le blocage des stimuli environnementaux potentielle-
ment interférents, les capacités stratégiques permettant
d’extraire des zones de stockage la trace mnésique utile à
un moment donné. Enfin, si l’on considère que les fonctions
exécutives ne sont pas purement cognitives mais aussi
sociales et affectives (voir ci-dessus), un déficit de l’admi-
nistrateur central de la mémoire de travail ne peut rendre
compte d’un défaut de prise en compte des conséquences
des actions, plutôt explicable par une diminution de la sen-
sibilité au renforcement ou d’un trouble de la cognition
sociale souvent dû à un défaut d’empathie, un émousse-
ment affectif ou un trouble de la théorie de l’esprit (partiel-
lement dépendant de l’administrateur central de la
mémoire de travail). Au total, un déficit isolé de l’adminis-
trateur central rend certainement compte d’une partie
importante du syndrome dysexécutif cognitif, mais il ne
peut expliquer l’ensemble « cognitif » de ce syndrome et
moins encore, le syndrome dysexécutif dans sa globalité si
l’on considère qu’il n’est pas impliqué dans la composante
affective et sociale des fonctions exécutives.
La psychologie expérimentale a permis d’isoler l’admi-
nistrateur central des autres composantes de la mémoire de
travail. La neuropsychologie permet d’aborder la question
de la relation entre un concept psychologique et son subs-
trat biologique. Dans le cas précis, existe-t-il une région ou
un réseau de régions cérébrales qui, lorsqu’elle ou il est
lésé(e), permet de rendre compte d’un déficit isolé de l’ad-
ministrateur central ? Le réseau cérébral dévolu à la
mémoire de travail est vaste et inclus des régions corticales
rétrorolandiques, des régions frontales et des structures
sous-corticales [6]. Au sein de ce vaste réseau, peut-on attri-
buer à l’une ou plusieurs structures le rôle spécifique d’ad-
ministrateur central de la mémoire de travail ?
La principale région candidate est la région latérale du
cortex préfrontal (CPFL ; Brodmann [BA], 8, 9, 10, 44, 45,
46 et 47) [6-9]. De haut en bas, le CPFL occupe la partie
latérale du gyrus frontal supérieur (BA 8/9), le gyrus frontal
moyen (BA 9/46), le gyrus frontal inférieur (BA 12/44/45).
Sa partie antérieure est représentée par la portion externe
du pôle frontal (BA 10). Ses afférences sont principalement
les régions associatives rétrorolandiques (les lobes parié-
taux et temporaux, y compris les régions internes hippo-
campiques, entorhinales et parahippocampiques), le CPFL
controlatéral, les autres régions frontales et la partie externe
(« parvocellulaire ») du noyau dorsomédian du thalamus
[6, 7]. Il projette principalement vers les structures motrices
du cerveau (les cortex prémoteur et moteur, les ganglions
de la base et les noyaux moteurs du tronc cérébral). Il réalise
donc une double interface entre, d’une part, les régions
d’intégration perceptive et les régions motrices [10] et,
d’autre part, entre les régions intégrant le passé (hippocam-
pes et régions associatives stockant les connaissances
sémantiques) et celles projetées vers l’action à venir (en par-
ticulier, les cortex prémoteurs qui vont programmer la
séquence d’actions à venir). Cette double interface est
essentielle pour assumer un rôle d’administrateur central.
En effet, l’administrateur central doit recevoir un signal
« montant » (bottom-up) des régions cérébrales où sont
stockées les représentations mentales (les régions associati-
ves rétrorolandiques) et dans le même temps exercer son
rôle exécutif (top down) en adressant un signal réciproque
vers ces mêmes régions (permettant l’adressage d’une
requête ou le filtrage attentionnel des stimuli non perti-
nents). Dans ce sens, les lésions du CPFL chez l’homme
augmentent précocement (20 ms après la stimulation),
l’amplitude des potentiels évoqués dans les régions senso-
rielles postrolandiques en réponse à des stimuli non perti-
nents, suggérant que le traitement sélectif des informations
pertinentes et non pertinentes est sous le contrôle inhibiteur
du cortex préfrontal [11]. Enfin, l’interface du CPFL vers des
régions telles que le cortex prémoteur, les ganglions de la
base et les noyaux moteurs du tronc cérébral lui permet de
jouer un rôle essentiel dans la préparation, l’exécution et le
contrôle de l’action.
Quels sont les arguments expérimentaux ou cliniques
permettant de relier le CPFL à l’administrateur central de
la mémoire de travail ? Les singes présentant une destruc-
tion bilatérale du CPFL ne peuvent effectuer correctement
une épreuve appelée la tâche des réponses différées
(delayed response task). Dans cette épreuve, l’animal est
soumis à un choix de réponse en fonction d’une informa-
tion présentée quelques secondes plus tôt mais séparée de
la réponse par un intervalle de plusieurs secondes appelé
« délai », et pendant lequel l’information n’est plus pré-
sente. Ainsi, ces épreuves découplent la perception de l’in-
formation de la réponse en interposant une phase tampon
qui a pour but d’obliger l’animal à sélectionner sa réponse
sur la base du stimulus maintenu sous la forme d’une repré-
sentation interne. De ce fait, au moment de sa réponse,
l’animal ne peut pas s’appuyer sur des indices externes
Article de synthèse
R
EVUE DE NEUROPSYCHOLOGIE
N
EUROSCIENCES COGNITIVES ET CLINIQUES
36
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

guidant son choix au moment de la réponse. Il doit donc
utiliser les données présentées avant le délai. Pour réaliser
cette tâche, le singe devra alors maintenir une trace (ou
représentation interne) de l’information pertinente et prépa-
rer mentalement l’action à venir. Il s’agit donc de tâches
« prototypiques » de mémoire de travail. Après une lésion
bilatérale du CPFL, le déficit est tellement sévère que le
comportement du singe paraît dicté par le hasard [12-16].
Plus précisément, au sein du CPFL, des lésions circonscrites
à sa partie moyenne, c’est-à-dire au cortex préfrontal dorso-
latéral englobant un sillon cortical, le sillon principal, suffi-
sent à produire un déficit massif [14, 17-21]. En revanche,
les lésions d’autres régions corticales associatives (frontales
ou plus postérieures) entraînent peu ou pas de déficit à cette
épreuve [12, 13, 15, 22]. Il faut bien noter que ces singes
frontolésés et présentant un déficit aux épreuves de répon-
ses différées n’ont pas de paralysie motrice ou de troubles
sensoriels primaires pouvant expliquer leur difficulté. Enfin,
lorsqu’ils effectuent cette épreuve en l’absence de délai, la
performance redevient normale, indiquant bien que le défi-
cit est lié à une difficulté à effectuer les opérations mentales
mises en jeu pendant le délai, c’est-à-dire la mémoire de
travail.
Toutefois, les données lésionnelles ci-dessus pourraient
suggérer, du moins chez le primate non humain, que le rôle
du CPFL est au moins aussi important pour le maintien en
mémoire à court terme que pour l’administrateur central. En
effet, dans les réponses spatiales différées, le choix se fait
entre deux stimuli. Pour répondre correctement, il suffit
donc de maintenir active la représentation du stimulus per-
tinent pendant le délai. Aucune manipulation de l’informa-
tion n’est utile. Ainsi, les lésions du CPFL chez le macaque
rhésus induisent un déficit qui peut être expliqué par un
défaut de maintien de l’information en mémoire à court
terme sans faire intervenir l’administrateur central [20].
Néanmoins, les données issues de l’enregistrement unitaire
de neurones du CPFL chez le singe pendant le délai des
réponses différées ont permis de mettre en évidence des
activités neuronales en relation avec diverses opérations
élémentaires typiquement dépendantes de l’administrateur
central. Outre les activités associées au maintien pendant
quelques secondes de l’information présentée sous la
forme d’une représentation mentale, suggérant que le
CPFL intervient dans le stockage en mémoire à court
terme (quelques secondes) [6, 7, 23], d’autres activités
sont en relation avec la préparation de l’action à venir
[24, 25], l’inhibition d’une action réflexive ou d’une infor-
mation non pertinente [24], au codage temporel d’un sti-
mulus dans une séquence [26] et à la sélection parmi plu-
sieurs réponses plausibles, suggérant leur implication dans
la construction de règles comportementales complexes
[27]. Au total, chez le primate non humain, il semble que
le CPFL joue un rôle essentiel pour le maintien en mémoire
à court terme, mais qu’il possède aussi, en termes d’activité
neurale, l’ensemble des outils permettant d’effectuer les
fonctions d’administrateur central. Ainsi, en suivant
Goldman-Rakic [28], le CPFL combinerait les fonctions
d’administrateur central et des systèmes tampons du
modèle de mémoire de travail de Baddeley.
Néanmoins, les activations du CPFL observées lorsque
le singe doit maintenir « passivement » l’information en
mémoire à court terme ne signifient pas nécessairement
que le maintien s’effectue dans le CPFL. En effet, le CPFL
est connecté de façon réciproque par des boucles réverbé-
rantes aux cortex associatifs rétrorolandiques, et il peut
envoyer vers ces régions un signal tonique durable permet-
tant le maintien à disposition de la représentation mentale
(sans transfert représentationnel vers le CPFL). Plusieurs
arguments plaident dans ce sens. Tout d’abord, lors des
tâches différées, des co-activations, en tout point identiques
à celles observées dans les neurones du CPFL, sont obser-
vées dans le cortex pariétal postérieur et le cortex temporal
inférieur avec lesquels le CPFL entretient des relations ana-
tomiques réciproques [29-34]. Ces observations indiquent
que la mémorisation à court terme est aussi effectuée dans
d’autres régions corticales en relations anatomiques direc-
tes avec le CPFL. De façon intéressante, Miller et Desimone
[35] ont comparé les profils d’activité des neurones dans
deux régions dans lesquelles les neurones ont la capacité
de maintenir une activité soutenue pendant la phase de
délai de tâches à réponses différées : le cortex inférotempo-
ral et le CPFL. Dans cette expérience, les singes voyaient
défiler une série de stimuli visuels séparés les uns des autres
par un bref délai. Ils avaient pour consigne de relâcher un
levier de pression quand un stimulus cible réapparaissait à
l’écran. Ce stimulus cible ne réapparaissait qu’après la pré-
sentation de plusieurs stimuli interférents. Des neurones
activés par le stimulus cible et maintenant leur activité de
façon soutenue pendant le délai étaient détectés dans le
cortex inférotemporal et le CPFL. Toutefois, dans le CPFL,
cette activité se maintenait jusqu’à la réapparition du stimu-
lus cible (et par conséquent jusqu’à la réponse comporte-
mentale), tandis que dans le cortex inférotemporal l’activité
cédait dès l’apparition du stimulus suivant. Ces données
suggèrent donc que dans le CPFL, l’activité de maintien
est dirigée vers la réponse comportementale, et qu’il ne
s’agit pas uniquement d’une activité liée à la mémorisation
per se. Chez l’homme, en imagerie fonctionnelle, Paulesu
et al. [36] ont montré que l’un des systèmes tampon de la
mémoire de travail, la boucle phonologique, était organisé
entre l’aire de Broca et une région du cortex pariétal posté-
rieur gauche, le gyrus supramarginal (BA 40). Cette dernière
région étant impliquée dans le maintien passif du matériel
verbal (phonological buffer), tandis que l’aire de Broca ali-
mente activement ce système passif de maintien à court
terme par la répétition sous-vocale. Ces données expéri-
mentales sont renforcées par la clinique. En effet, une
forme clinique de la maladie d’Alzheimer à début posté-
rieur, appelée aphasie progressive logopénique ou phono-
logique [37], commençant par l’atteinte symptomatique du
lobule pariétal postérieur gauche se manifeste initialement
par un trouble majeur de la boucle phonologique altérant
les capacités de maintien en mémoire de travail verbale et
se répercutant sur les capacités de compréhension et de
Article de synthèse
R
EVUE DE NEUROPSYCHOLOGIE
N
EUROSCIENCES COGNITIVES ET CLINIQUES
37
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

répétition de mots longs (c’est-à-dire comprenant un nom-
bre important de syllabes).
Ainsi, même si le CPFL intervient dans le maintien sim-
ple de l’information en mémoire de travail, son rôle critique
pourrait se trouver ailleurs, en particulier dans les tâches de
gestion exécutive (ou d’administrateur central). En particu-
lier, un des rôles principaux résiderait dans l’utilisation des
représentations mentales pour préparer l’action volontaire à
venir. Plusieurs études d’imagerie fonctionnelle chez
l’homme sain ont montré des activations significativement
plus importantes dans le CPFL au cours de tâches nécessi-
tant une préparation plus intense de l’action à venir (telle
qu’une version automatisée de l’épreuve de la « Tour de
Londres ») par comparaison à des épreuves dans lesquelles
seul le maintien « passif » en mémoire de travail était requis
[38-40]. Dans le même ordre d’idée, dans notre laboratoire,
nous avons montré en IRMf à l’aide de tâches dérivées des
réponses différées chez le singe (les épreuves dites de
« séquences spatiales mémorisées » et de « double délai »)
que l’activation du CPFL dépend au premier chef de l’utili-
sation mentale de l’information maintenue en représenta-
tion mentale pour préparer l’action à venir et peu ou pas
du simple maintien en mémoire de travail [41, 42]. Le sim-
ple maintien en mémoire à court terme n’active pas le
CPFL. En revanche, lorsque le maintien en mémoire de tra-
vail requiert la mise en jeu de stratégies exécutives comme
le classement temporel, alors le CPFL est activé [42]. Au
total, l’ensemble de ces travaux souligne l’implication de
la région latérale du CPF dans la gestion exécutive (mani-
pulation et préparation) d’un comportement élaboré.
Quelle est l’organisation anatomo-fonctionnelle du
CPFL pour mettre en place ses fonctions d’administrateur
central de la mémoire de travail ? Les travaux chez l’animal
et l’homme suggèrent qu’il existe, parallèlement à l’organi-
sation modulaire cognitive décrite par Baddeley, une orga-
nisation modulaire anatomique de la mémoire de travail,
dans laquelle différentes régions cérébrales sont impli-
quées, plus ou moins spécifiquement, dans un ou plusieurs
des processus cognitifs décrits dans le modèle de Baddeley.
Deux axes principaux de différenciation du rôle de ces dif-
férentes régions dans la mémoire de travail guident les étu-
des anatomo-fonctionnelles. Premièrement, une différen-
ciation en fonction du domaine de l’information traitée
(par exemple spatial, verbal ou figuratif dans la modalité
visuelle) ; secondairement, une différenciation en fonction
des processus cognitifs impliqués par la tâche étudiée (par
exemple, opération de stockage à court terme, de compa-
raison ou de manipulation active des informations en
mémoire de travail).
L’organisation du cortex rétro-rolandique fait apparaître
une ségrégation fonctionnelle principalement domaine-
dépendante. La mémoire de travail visuo-verbale implique-
rait principalement le cortex pariétal inférieur gauche (gyrus
supramarginal gauche) [36, 43, 44]. La mémoire de travail
visuo-spatiale paraît impliquer de façon prédominante le
cortex pariétal droit [43, 45, 46]. Plus précisément, les cor-
tex pariétaux droits inférieur (gyrus supramarginal, BA 40)
et supérieur (BA 7) paraissent le plus souvent impliqués
dans les tâches de mémoire de travail visuo-spatiale [47].
Quant à la mémoire de travail visuelle non spatiale (stimuli
de type objets ou images abstraites, visages), elle ferait
intervenir de façon prédominante le cortex pariétal posté-
rieur gauche mais aussi le cortex temporal inférieur gauche
[47, 48]. Ainsi, en modalité visuelle, il semble exister une
ségrégation domaine-dépendante épousant la dissociation
dorsoventrale décrite par Mishkin et Ungerleider dans le
traitement des informations visuelles par le cortex rétro-
rolandique [49]. La latéralisation interhémisphérique sug-
gérée par certaines de ces études n’est évidemment pas
absolue, apparaissant principalement lorsque l’on compare
directement les domaines verbal et spatial entre eux. Le rôle
du cortex pariétal gauche (gyrus supramarginal) a été spé-
cifiquement étudié en mémoire de travail verbale [36, 44,
47]. Cette région semble impliquée dans le stockage passif
des informations verbales. Le cortex pariétal supérieur droit
serait impliqué dans la récapitulation des informations spa-
tiales, impliquant des déplacements d’attention spatiale
focale [47, 50-52], en particulier dans le stockage passif à
court terme [47, 48]. Les régions rétrorolandiques semblent
donc suivre une organisation en fonction du domaine de
l’information visuelle traitée et être impliquées plus spécifi-
quement dans les processus de maintien en mémoire à
court terme en mémoire de travail.
En avant du sillon rolandique, les modèles actuellement
proposés d’organisation anatomique et fonctionnelle du
CPFL suggèrent que celui-ci peut aussi être divisé en sous-
régions selon la nature des processus cognitifs mis en jeu.
Plusieurs modèles sont proposés à partir des données
expérimentales :
•les travaux de Fuster [7, 53] et plus récemment de Miller
et al. [54-56] chez le singe et de D’Esposito et al. chez
l’homme [57] plaident pour un rôle d’intégration supramo-
dale du CPFL dont la fonction serait dirigée vers les aspects
les plus exécutifs de la mémoire de travail. Dans ce modèle,
le CPFL n’est pas divisible en plusieurs sous-régions effec-
tuant un traitement différent sur l’information maintenue en
mémoire de travail. Au contraire, cette fonction de gestion
des ressources mentales nécessite l’intégration d’informa-
tions de nature sensorielle multiple ;
•pour Petrides et Owen, le CPFL est organisé en deux sous-
régions (inférieure et supérieure) différentes pour la nature
des opérations effectuées en mémoire de travail [58-61].
Dans ce modèle à « deux étages », la région supérieure
(BA 9 et la partie supérieure de BA 46) est impliquée dans
la manipulation et la gestion séquentielle des informations
maintenues en mémoire de travail. La région inférieure (BA
12/45/47 et la partie inférieure de BA 46) assurerait le main-
tien en mémoire de travail et la réalisation de comporte-
ment nécessitant peu ou pas de manipulation mentale (par
exemple, juger si une information maintenue en mémoire
de travail doit être manipulée ou non) ;
•Goldman-Rakic et al. soutiennent un modèle différent de
ségrégation du CPFL. Dans ce modèle, le CPFL est subdi-
visé en modules anatomiques de mémoire de travail diffé-
Article de synthèse
R
EVUE DE NEUROPSYCHOLOGIE
N
EUROSCIENCES COGNITIVES ET CLINIQUES
38
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.
 6
7
8
6
7
8
1
/
8
100%