chentouf hanane - Université d`Oran 1 Ahmed Ben Bella

ﺔـــــــــــﯿــــــﺒــﻌـﺸﻟا ﺔـــــــﯿطاﺮﻘﻤﯾﺪﻟا ﺔﯾﺮـــﺋاﺰﺠﻟا ﺔﯾرﻮــــﮭﻤﺠﻟا
République Algérienne démocratique et populaire
Ministère de l’enseignement supérieur et de la recherche scientifique
Université d’Oran Ahmed Ben Bela -1-
Faculté des sciences
Département de biologie
Thèse de doctorat (LMD)
Spécialité : Microbiologie appliquée
Option : Contrôle microbiologique et hygiène alimentaire
Thème :
Effet des substances antimicrobiennes produites
par Leuconostoc mesenteroides isolées à partir
du lait de chamelle Algérien sur Listeria spp.
dans les produits alimentaires
Présenté par : MIMOUN née CHENTOUF Hanane Fatma
Devant les membres du jury :
Président :
Mr KIHAL Mebrouk
Professeur (Université d’Oran)
Examinateur :
Mr ABBOUNI Bouziane
Professeur (Université de Sidi Bel Abbes)
Examinateur :
Mr BABA HAMED Mohammed Bey
Professeur (Université d’Oran)
Examinateur :
Mr HADADJI Miloud
Professeur (Université d’Oran)
Rapporteur :
Mlle BENMECHERNENE Zineb
M. C. A. (Université d’Oran)
2014-2015

Remerciements
Le premier de mes remerciements ira au bon Dieu miséricorde qui m’a donné la santé,
la volonté et la patience pour réaliser ce modeste travail.
Ce travail n’aurait vu le jour sans la confiance et surtout la patience de ma directrice de
recherche Madame BENMECHERNENE Zineb qui m'a bien accueillie pendant ces années
de thèse. Je tiens à lui exprimer toute ma reconnaissance d'avoir su m'encadrer, me conseiller
et me soutenir tout en me laissant travailler très librement et surtout d’avoir était une bonne
amie.
Mes plus sincères remerciements vont à Monsieur le Professeur KIHAL Mebrouk qui
m'a fait l'honneur d'accepter de juger mon travail et dont l'enseignement et la passion de ce
dernier resteront des exemples pour moi.
Je remercie Messieurs les Professeurs ABBOUNI Bouziane, BABA HAMED
Mohamed Bey et HADADJI Miloud qui ont eu la gentillesse d'accepter de juger mon
travail.
Je tiens à exprimer ma reconnaissance à Melle ZAROUR Kanza qui m'a aidé,
encouragé et soutenue dans mes moments d'enthousiasme étincelant comme dans les
moments de doute inconsolables.
Je tiens à remercier vivement Monsieur BARROS-VELAZQUEZ Jorge qui m’a
accueillie au sein de son laboratoire de microbiologie à l’université de Santiago De
Compostella de Lugo (Espagne), de me faire partie de son équipe et de payer mon stage sans
hésitation et pour m'avoir accordé sa confiance en m'offrant la possibilité de poursuivre ce
travail.
Toutes mes gratitudes vont à Messieurs QUINTELA BALUJA Marcos et El Nakib
Mohamed de leur patience et de m’avoir énormément aidé.
Je remercie tous qui m’ont aidé de près ou de loin afin d’accomplir ce modeste travail.

Dédicace
Je dédie ce modeste travail
A mon très cher père Benali qui m'a toujours soutenu, et a été toujours présent
pour moi.
A la plus chère au monde, ma mère Turkia qui m’a toujours encouragé et me
réconforter durant mes études.
A mon adorable cher mari Djamel Eddine qui m’a supporté et d’avoir été
très compréhensible.
A mon petit bout de chou Mohamed Aymene
A la mémoire de ma sœur Imane (Allah yarhamha)
A mes frères Mohamed Yacine et Sofiane
A ma belle-famille : Fadéla, Houari, Wafaa, Belkhir, Mokhtaria et Tarek
A toute ma famille
A toutes mes amies.

1
Table des matières
Problématique
ﺺﺨﻠﻣ
Abstract
Résumé
Liste des abréviations
Liste des figures
Liste des tableaux
Introduction ................................................................................................................................ 1
Synthèse bibliographique
1. Généralités sur le dromadaire ............................................................................................. 3
1.1. Origine du dromadaire ................................................................................................ 3
1.2. Taxonomie et race ....................................................................................................... 3
1.3. Répartition géographique des dromadaires ................................................................. 4
1.3.1. Distribution dans le monde ......................................................................................... 4
1.3.2. Distribution en Afrique ............................................................................................... 5
1.3.3. Distribution en Algérie ............................................................................................... 6
1.4. Dromadaire à vocation lait .......................................................................................... 6
1.4.1. Production laitière mondiale ....................................................................................... 7
1.4.2. Production laitière en Algérie ..................................................................................... 7
2. Généralités sur le lait........................................................................................................... 8
2.1. Lait de chamelle .......................................................................................................... 9
2.1.1. Caractéristiques organoleptiques et physicochimiques du lait camelin ...................... 9
2.1.2. Propriétés nutritionnelles du lait camelin.................................................................... 9
2.1.3. Caractéristiques microbiologiques du lait camelin ................................................... 10
2.1.4. Système protecteur du lait camelin ........................................................................... 10
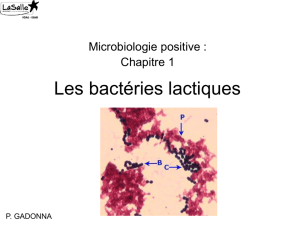
3. Les bactéries lactiques ...................................................................................................... 11
3.1. Habitat et origine des bactéries lactiques .................................................................. 11
3.2. Taxonomie des bactéries lactiques ............................................................................ 11
3.3. Caractéristiques des principaux genres des bactéries lactiques ................................ 13
3.3.1. Le genre Lactobacillus .............................................................................................. 13

2
3.3.2. Le genre Lactococcus................................................................................................ 13
3.3.3. Le genre Streptococcus ............................................................................................. 14
3.3.4. Le genre Enterococcus .............................................................................................. 14
3.3.5. Les genres Leuconostoc, Oenococcus et Weissella .................................................. 14
3.3.6. Les genres Pediococcus et Tetragenococcus ............................................................ 15
3.3.7. Le genre Bifidobacterium.......................................................................................... 15
3.4. Les propriétés antimicrobiennes des bactéries lactiques........................................... 16
3.4.1. Le pH et les acides organiques .................................................................................. 16
3.4.2. Le peroxyde d’hydrogène ......................................................................................... 17
3.4.3. Le dioxyde de carbone .............................................................................................. 17
3.4.4. Le diacétyl ................................................................................................................. 18
3.4.5. La reutérine ............................................................................................................... 18
3.4.6. Les bactériocines ....................................................................................................... 18
4. Le genre Leuconostoc ....................................................................................................... 19
4.1. Historique .................................................................................................................. 19
4.2. Habitat ....................................................................................................................... 19
4.3. Classification ............................................................................................................. 21
4.4. Caractérisation des Leuconostoc ............................................................................... 23
4.5. Le pouvoir antimicrobien des Leuconostoc .............................................................. 24
4.6. Résistances aux conditions de stress ......................................................................... 25
4.7. Utilisation des souches de Leuconostoc dans l’industrie alimentaire ....................... 26
5. Les bactériocines ............................................................................................................... 26
5.1. Historique .................................................................................................................. 26
5.2. Définition .................................................................................................................. 27
5.3. Classification ............................................................................................................. 28
5.3.1. Classe I : Les lantibiotiques ...................................................................................... 28
5.3.2. Classe II .................................................................................................................... 28
5.3.3. Classe III ................................................................................................................... 30
5.3.4. Classe IV ................................................................................................................... 31
5.4. La production des bactériocines ................................................................................ 32
5.5. Le conditionnement des bactériocines ...................................................................... 33
5.6. L’application des bactériocines et la bactérie productrice dans le secteur alimentaire
33
5.6.1. Application de la bactériocine dans le secteur alimentaire ....................................... 33
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
1
/
190
100%