Corrigé DC3 4sc LFH2011 - E

1
Lycée Farhat Hached M’saken Correction du devoir de contrôle n°3
Section : Sciences Expérimentales Classe : 4sc.1
Epreuve : Sciences de la Vie et de la Terre Année scolaire : 2010-2011
Corrigé Barème
PREMIERE PARTIE : (12 points)
QCM : (6 points)
1
2
3
4
5
6
c
a
c, d
a
b
a
Toute réponse fausse annule la note attribuée à l’item.
Immunité (6 points)
Les réactions immunitaires spécifiques impliquent la capacité des cellules immunocompétentes à
distinguer le soi du non-soi. On appelle soi l’ensemble des molécules résultant de l’expression du
génome d’un individu et non-soi les molécules qui ne sont pas normalement codées par le
génome et sont reconnues comme étrangères par l’organisme.
I- La reconnaissance du non-soi
Les cellules chargées de reconnaître le non-soi sont des lymphocytes. Dans le cas d’une réponse à
médiation humorale, il s’agit de lymphocytes B (LB), cellules qui acquièrent leur
immunocompétence dans la moelle osseuse. Les LB sont capables de reconnaître des antigènes
circulants comme ceux portés par certains microorganismes. Cette capacité est liée à la présence
de récepteurs membranaires, les récepteurs B, capables de reconnaître et de lier un antigène de
façon spécifique, chaque clone de LB ne reconnaissant qu’un seul antigène car il ne possède
qu’un seul type de récepteur B. Les récepteurs B sont des anticorps membranaires(Ig). Ils sont
constitués de quatre chaînes polypeptidiques identiques deux à deux, deux chaînes lourdes et
deux chaînes légères. Chaque chaîne comporte une région constante et une région variable, les
régions constantes des chaînes lourdes permettant la fixation à la membrane de la cellule et les
régions variables étant responsables de la reconnaissance et de la liaison de l’antigène.
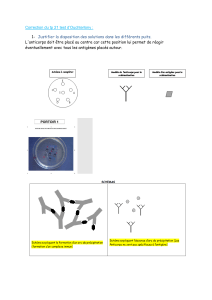
Le schéma de la structure générale
d’un anticorps :
Lorsque la molécule antigénique
présente une configuration spatiale
complémentaire de celle du site
correspondant de la molécule
d’anticorps (site Ac ou paratope),
elles se lient entre elles formant un
complexe-immun qui permet la
neutralisation de l’antigène sans
pouvoir l’éliminer.
1 x 6
0,5
1
1,5
6
points
6
points

2
II- La neutralisation du non-soi
La reconnaissance d’un antigène par un LB permet la sélection d’un clone spécifique par
l’activation du LB qui se multiplie et donne naissance à des plasmocytes, cellules productrices
d’anticorps circulants, les immunoglobulines. Il faut noter que, le plus souvent, l’activation de la
réponse immune nécessite la coopération de lymphocytes T4 auxiliaires qui stimulent les LB par
l’IL2, messager chimique intercellulaire. Les anticorps sécrétés par les plasmocytes possèdent la
même structure que les anticorps membranaires mais ils circulent en solution dans le sang. Ils
reconnaissent le même déterminant antigénique que celui à l’origine de la stimulation du LB et se
lient à lui recouvrant de molécules d’anticorps le microorganisme qui le porte. Du fait de leur
double site antigène, les anticorps peuvent aussi constituer des complexes immuns qui immobilise
les microorganismes.
III- L’élimination du non-soi
Les molécules d’immunoglobulines circulantes comportent un site de liaison pour le complément.
Le complément est un ensemble de protéines non spécifiques de l’antigène qui s’activent en
cascade lorsque les immunoglobulines se lient aux antigènes. Cette cascade aboutit à la formation
d’un complexe lytique (CAM)détruisant la cellule cible. Les déchets sont alors éliminés par
phagocytose. Les anticorps fixés au non-soi, par exemple une bactérie, peuvent aussi être
éliminés directement par phagocytose car la partie constante des immunoglobulines est reconnue
par un récepteur spécifique des cellules phagocytaires, macrophages et granulocytes . Les
complexes immuns sont également phagocytés et détruits par les macrophages et les
granulocytes.
Conclusion
La réponse immunitaire spécifique à médiation humorale illustre les principales caractéristiques
des réponses immunitaires spécifiques : spécificité, intervention de cellules spécialisées
fabriquant des molécules spécifiques, membranaires et solubles, coopération intercellulaire,
intervention des LT4, même si elle paraît moins essentielle qu’au cours de la réponse à médiation
cellulaire.
Deuxième partie :(8 points):
Le fonctionnement du système nerveux central repose sur les échanges de messages nerveux entre
neurones au sein de réseaux neuronaux. Les documents proposés permettent de proposer une
explication au mécanisme de la transmission des messages d’un neurone à un autre.
Le document 4 montre qu’une stimulation électrique de l’axone de N1 donne naissance à un
message nerveux, représenté ici par un potentiel d’action. On constate qu’un message similaire
est transmis à N2. La zone de contact entre N1 et N2 photographiée sur le document 3b est une
synapse. La région présynaptique située à gauche du cliché correspond au neurone N1 et contient
des vésicules synaptiques à acétylcholine. La région post synaptique, correspondant au neurone
N2 est située à droite du cliché et est séparée de la précédente par un espace synaptique. Dans ces
conditions, un message électrique ne peut franchir la synapse. Le document 4 montre l’action de
l’acétylcholine sur la membrane post synaptique (B) : à la suite du dépôt d’une faible
concentration d’acétylcholine dans la synapse, on constate une faible dépolarisation de la
membrane post synaptique. Cette substance est donc capable de modifier le potentiel de
membrane du neurone N2. Toutefois, la dépolarisation reste locale, contrairement à l’expérience
A et aucun potentiel d’action n’est enregistré en R3. En revanche, lorsque la concentration en
acétylcholine est plus élevée, on constate que la dépolarisation de la membrane post synaptique
est plus élevée et engendre un potentiel d’action en R2 (C). De plus, ce potentiel se propage
1,5
1
0, 5
0,5
1
1
1

3
puisqu’on l’enregistre également en R3. Enfin, la forme du tracé obtenu en R2 dans cette
expérience est identique à celle obtenue en R2 dans l’expérience A. Étant donné qu’il existe des
vésicules à acétylcholine dans la terminaison présynaptique, on peut penser que la stimulation du
neurone N1 conduit à la libération d’acétylcholine dans l’espace synaptique.
L’action de l’acétylcholine ne se produit que lorsqu’on l’injecte dans l’espace synaptique : son
injection dans N2 ne produit aucun effet. Elle interagit donc avec la face extracellulaire de la
membrane. Étant donné qu’elle y produit une dépolarisation d’autant plus élevée qu’elle est plus
concentrée (B et C), elle est en mesure de modifier les flux ioniques à travers la membrane. Elle
agit donc sur des canaux ioniques chimiodépendants.
Enfin, l’effet de la pilocarpine permet d’expliquer l’action brève de l’acétylcholine : dans les
conditions physiologiques, l’acétylcholinestérase hydrolyse l’acétylcholine dans la fente
synaptique dès qu’elle a agi sur ses récepteurs. En présence de pilocarpine, inhibiteur de
l’enzyme, l’acétylcholine n’est pas hydrolysée et son action est prolongée.
Le schéma suivant résume ce mécanisme de transmission chimique responsable du
fonctionnement synaptique et, donc, de la transmission des messages entre neurones.
1
1
1
1,5
1
/
3
100%