Propriétés mécaniques des cellules et tissus - ENS-phys

Manon Michel Rapport du séminaire
L3 de physique du 9 décembre 2008
Compte-rendu du séminaire de la FIP du 9 décembre 2008
Propriétés mécaniques des cellules et tissus
Jean-François Joanny
Institut Curie
Introduction
I. Cytosquelette Actine-Myosine et systèmes actifs
1. Présentation du cytosquelette
2. Expériences
3. Structure d'un gel acto-myosinique
4. Systèmes actifs
II. Théorie hydrodynamiques des gels actifs
1. Théorie hydrodynamique
2. Contrainte mécanique
III. Anneaux contractiles
1. Pinçage d'anneau dans des oeufs de Xénope (étudié par Mandato,
Bement)
2.Théorie des gels actifs appliquée aux anneaux contractiles
3. Formation d'un anneau pendant la mitose
Conclusion
Questions de la salle
1

Manon Michel Rapport du séminaire
L3 de physique du 9 décembre 2008
Introduction
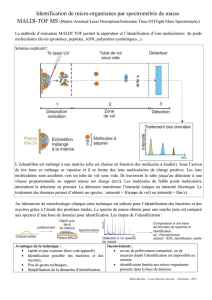
Le monde cellulaire est un univers très dynamique. Les cellules se divisent, se déplacent,
échangent des informations entre elles… Toutes ces fonctions indispensables à la vie cellulaire
nécessitent de l’énergie. La source d’énergie des cellules est l’ATP, une petite molécule comportant
trois phosphates. Ce sont les liaisons chimiques entre les phosphates qui, lorsqu’elles se rompent,
fournissent l’énergie aux cellules. La cellule peut alors utiliser cette énergie chimique et la
transformer en énergie mécanique grâce aux moteurs moléculaires.
Jean-François Joanny, chercheur à l'institut Curie, a présenté une approche quantitative et
physique du fonctionnement des moteurs moléculaires en modélisant le cytosquelette comme un
système hydrodynamique qui ne serait pas à l'équilibre thermodynamique (dit système actif).
Par manque de temps, seule la partie sur le comportement actif des cellules a été exposée.
Molcule d'ATP
2

Manon Michel Rapport du séminaire
L3 de physique du 9 décembre 2008
I. Cytosquelette Actine-Myosine et systèmes actifs
1. Présentation du cytosquelette
Le cytosquelette d'une cellule est l'ensemble organisé des polymères biologiques qui lui
confèrent l'essentiel de ses propriétés mécaniques. La référence terminologique au « squelette » des
vertébrés est cependant trompeuse puisque :
–toutes les composantes du cytosquelette sont renouvelées par polymérisation en
permanence.
–Le cytosquelette est à l'origine de la plupart des forces exercées par la cellule pour se
déplacer et se nourrir, ce en quoi il s'apparente plutôt à un ensemble de « muscles ».
–enfin, les propriétés mécaniques du cytosquelette sont très variables suivant les composantes
et les situations considérées.
Le cytosquelette des eucaryotes est constitué de polymères biologies protéiques, que l'on
nomme fibres au vu de leur grande taille à l'échelle cellulaire. Ces polymères sont classés en trois
catégories : les filaments d'actine (que l'on retrouve également dans les fibres musculaires) d'un
diamètre de 5 à 9 nm, les filaments intermédiaires d'un diamètre de 10 nm et les microtubules d'un
diamètre de 25 nm. Les polymères sont organisés en réseaux, faisceaux ou câbles suivant les rôles
qu'ils remplissent. Ce haut niveau d'organisation est rendu possible par la présence de centaines de
protéines auxiliaires (protéines de pontage, de branchement, protéines capuchon et
dépolymérisantes, d'ancrages).
Durant le séminaire, il n'a été question que des filaments d'actine et des moteurs
moléculaires constitués de myosine.
Eléments du cytosquelette
(Bleu : noyaux, Vert : microtubules, Rouge : actine)
3

Manon Michel Rapport du séminaire
L3 de physique du 9 décembre 2008
2. Expériences
Avant de rentrer dans les détails de la constitution des filaments d'actine et des moteurs
moléculaires, voici les résultats de deux expériences qui mettent en lumière le lien entre ces derniers
et les propriétés mécaniques d'une cellule.
Première expérience :
On place dans un tube capillaire d'un diamètre de 400 nm et accéléré 180 fois un gel d'
actine-myosine. A t = 0 s, on libère de l'ATP, ce qui revient à libérer de l'énergie dans le
système.
On observe alors le ménisque se creuser. En effet, les moteurs moléculaires créent une
tension dès la libération de l'énergie, tension qui amène à une contraction plus forte du
gel.
Connaissant la courbure du ménisque et les tensions superficielles de surface, on peut
alors calculer la pression et les contraintes subies par le système, qui sont de l'ordre de
1000 Pa.
(Expérience de G. Koenderink et D. Cuvelier)
Gel dans le capillaire
Deuxième expérience :
On empêche une cellule (fibroplaste T3) de s'accrocher à une
surface, des ions calcium sont présents dans le milieu
extérieur.
On observe alors des oscillations stables (elles ne cessent
qu'au bout de plusieurs heures). La période de ces oscillations
est extrêmement bien définie (de l'ordre de 30s), même si une
dérive est observée au bout d'un temps long.
Fibroplaste T3
Il est intéressant d'observer les réponses de ce système lors de l'injection de différentes
drogues. Ainsi, si une drogue qui détruit l'actine est libérée dans le milieu, les oscillations cessent,
de même si on libère une drogue qui détruit les moteurs moléculaires.
4

Manon Michel Rapport du séminaire
L3 de physique du 9 décembre 2008
La présence du calcium dans le milieu extérieur est importante. La membrane de la cellule
possède des canaux, par lesquels le calcium va pénétrer, les oscillations ne commencent que suite à
cette pénétration et si la concentration en calcium est supérieure à un certain seuil. Ainsi, une fois
les canaux bouchés, les oscillations cessent.
Ainsi, les oscillations dépendent des propiétés
contractiles de l'actine et de la concentation en
calcium (via un seuil).
La période des oscillations d'environ 30s
dépend de l'activité de la myosine (ie les
moteurs moléculaires).
Oscillations du rayon
Les oscillations s' interprètent comme une instabilité mécanique de la couche corticale de la
cellule (là où l'actine est localisée).
(Expérience de P. Pullarkat)
On a réussi à modéliser ces phénomènes de façon satisfaisantes.
3. Structure d'un gel acto-myosinique
•Les Filaments d'actine
Les filaments sont polaires, du fait de l'asymétrie du monomère d'actine et de leur
assemblage en hélice, et sont l'objet de « treadmilling » (tapis roulant en français), ie une des
extrémités (dite (+)) peut polymériser beaucoup plus vite que l'autre (dite (-)), il en résulte un
déplacement d'équilibre vers la polymérisation du côté (+) et vers la dépolymérisation du côté (-).
Ces deux déplacements d'équilibre concomitants font que la chaîne croît en permanence du côté (+)
et décroît du côté (-).
5
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%