Stabilite de la determination et de la differentiation

/.
EmbryoL
exp.
Morph.
Vol.
36,
1,
pp.
183-196,
1976
183
Printed
in Great
Britain
Stabilite
de la
determination
et de
la differentiation
des
caracteres sexuels somatiques
chez Nereis pelagica
L.
(Annelide Polychete)
ParYOLANDE BOILLY-MARER1
Universite des Sciences
et
Techniques de Lille
I,
France
SUMMARY
Stability of
determination and differentiation
of the
somatic
sexual
characters
in
Nereis pelagica
L.
{Annelida polychaeta)
Experiments
of
regeneration
in
situ
or on
grafts were carried
out to
test
the
stability
of
determination
of
the somatic sexual characters
in
Nereis
pelagica.
The sexual characters
of
the parapodial cirri (male
and
female swellings, male crenella-
tions)
are
always expressed
on
stump
or
regenerate according
to
the
genetic
sex.
Super-
numary parapodia which developed
at the
site of junction
of
the heterologous faces
of
host
and graft
of
opposite sexes, might have
the
characters
of
either sex; this result
can be
inter-
preted
by
supposing that
a
supernumary parapodium
is
derived from tissues
of
either host
or graft,
of
the corresponding sex.
Conversely,
the
pygidial papillae, which normally occur only
on the
heteronereid male,
will appear
on
regenerating female pygidia, triggered
to
transform
to the
heteronereid
con-
dition naturally
or
by decerebration, with a frequency depending on the age
of
the
regenerate,
the length
of
the regenerate,
and
the genital condition
of
both the stump
and
the regenerate.
It
is
concluded that
the
presence
of
the papillae is
not a
sex-specific character,
but
that their
development
is
normally inhibited
in the
maturing female.
INTRODUCTION
Chez les Nereidiens
a
epitoquie,
le
dimorphisme sexuel
se
met
en
place
au
cours
de
l'heteronereidation
au
niveau des cirres parapodiaux
et
du pygidium.
Les Nereis pelagica L. males se caracterisent essentiellement par
la
presence
de
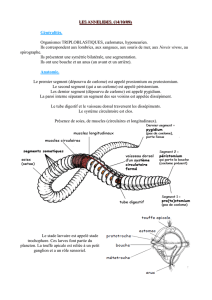
crenelures sur les cirres parapodiaux dorsaux de la region posterieure (Fig. 1)
et
de papilles pygidiales. D'autre part, les cirres deviennent renfles sur un nombre
de parapodes plus important chez
le
male (sept premieres paires) que chez
la
femelle (cinq premieres paires) (Fig. 2).
Ces caracteres sont determines precocement avant
la
differenciation sexuelle,
soit uniquement chez le male (crenelures), soit
a
la fois chez le male et la femelle
(renflement des cirres des cinq premieres paires de parapodes, papilles pygidiales)
(Boilly-Marer, 1974a). Toutefois,
la
differenciation des papilles pygidiales n'est
1
Adresse
de
Vauteur:
Universite des Sciences
et
Techniques de Lille
I,
Service de Biologie
Animale,
L.A.
au
C.N.R.S.
n°
148
(Endocrinologie comparee
des
Invertebres),
B.P. 36,
59650 Yilleneuve d'Ascq, France.

184YOLANDE BOILLY-MARER
FIGURES 1-2
Transformation heteronereidienne des cirres parapodiaux chez N.
pelagica
L.
Fig. 1. Cirres parapodiaux dorsaux de la region posterieure. Heteronereis male. Des
crenelures (fleches) apparaissent dans la zone moyenne du cirre. Les zones apicale et
basale en sont depourvues.
Fig. 2. Parapodes de la region anterieure. Heteronereis male. Les cirres dorsaux (cd)
et ventraux (cv) presentent un renflement (fleches) dans la zone moyenne. La zone
apicale du cirre n'est pas transformee.
realisee, dans les conditions normales, que chez le male; ce caractere peut
cependant s'exprimer chez la femelle si celle-ci subit l'epitoquie experimentale a
l'etat jeune ou en presence d'un milieu male obtenu par injection du contenu
coelomique male ou greffe sur un male (Boilly-Marer, 1914b).
Nous nous proposons ici d'apprecier le degre de stabilite de ces caracteres
sexuels somatiques en provoquant leur differentiation sur des cirres para-
podiaux et pygidiums regeneres.
MATERIEL ET METHODES
Toutes les experiences ont ete realisees sur des N. pelagica L. d'age varie,
sexuellement differenciees ou non. Les femelles ont ete reparties en six lots
suivant le diametre ovocytaire (20-30
jam,
40 /im, 50-60
ju,m,
70-80
jLtm,
90-100 /on, 130 /mi) mesure au debut de l'experience (diametre de l'ovocyte
mature: 200/im environ). La regeneration des structures sur lesquelles se

La differentiation des
caracteres
sexuels
somatiques
chez Nereis 185
developpent les caracteres sexuels somatiques (cirres parapodiaux et pygidium)
est provoquee soit par ablation elective des cirres parapodiaux ou des parapodes,
soit par amputation caudale de 10 a 35 segments. Dans ce dernier cas, l'etat
genital du regenerat est precise par l'etude des coupes histologiques.
D'autre part, la neoformation de parapodes surnumeraires est induite suivant
le procede decrit dans un travail anterieur (Boilly-Marer, 1969 et
1971
a et b) par
juxtaposition de territoires heterologues (dorsal
<-•
ventral) a l'aide de greffes de
paroi de corps.
L'etude de la regeneration (pygidium + parapodes) est faite soit sur une
Nereis, soit sur un greffon de sexe donne place sur un porte-greffe de sexe oppose.
Lorsque l'epitoquie naturelle ne se produit pas au cours de F experience, la
differentiation des caracteres sexuels somatiques est provoquee en declenchant
l'heteronereidation par decerebration, soit au moment de l'amputation, soit
8 jours a 6 mois apres celle-ci. La nature des caracteres sexuels est analysee au
moment ou l'epitoquie des Nereis est atteinte, c'est-a-dire 30 a 35 jours apres la
decerebration.
RESULTATS
Nous envisagerons successivement revolution des cirres parapodiaux
regeneres, puis celle des pygidiums neoformes.
A. Evolution des
cirres
parapodiaux regeneres
Nous distinguerons les cirres des parapodes edifies apres amputation de la
partie posterieure du corps, des parapodes, ou des cirres parapodiaux eux-
memes, puis ceux des parapodes induits par juxtaposition de territoires hetero-
logues (parapodes surnumeraires).
(1) Parapodes regeneres
Dans tous les cas, les organes amputes (segments, parapodes, cirres para-
podiaux) regenerent dans un delai de 2 a 6 mois qu'il s'agisse de regeneration
in situ (a) ou sur le greffon (b).
(a) Regeneration in situ
Lorsque l'ablation des structures portant les cirres parapodiaux ou des cirres
parapodiaux eux-memes est pratiquee sur des Nereis atoques, devolution des
cirres parapodiaux regeneres au cours de l'heteronereidation (43 cas) est toujours
identique a celle des cirres parapodiaux voisins n'ayant pas subi l'amputation,
et ceci meme si la regeneration du cirre est incomplete (moitie a deux tiers de la
longueur initiale du cirre).
Lorsque l'amputation des cirres parapodiaux dorsaux ou des parapodes
posterieurs est effectuee sur des Nereis sub-epitoques (st. 1 de la terminologie
de Durchon, 1952) et jusqu'a 20 jours environ avant le stade final de l'hetero-
nereidation, les cirres parapodiaux dorsaux sont d'abord regeneres sous leur

186YOLANDE BOILLY-MARER
FIGURES 3-4
Homogreffe en position caudale d'un troncon de corps femelle sur un
porte-greffe male (N.
pelagica).
Fig. 3. Deux mois apres la greffe; immediatement apres la decerebration. La suture
entre porte-greffe et greffon est realisee (fleche); ce dernier a regenere huit segments
setigeres.
Fig. 4. Meme
Nereis
que sur Fig. 3,
1
mois apres la decerebration (soit
3
mois apres
la greffe). Le porte-greffe et le greffon sont epitoques. Remarquer la transformation
moins poussee des parapodes du greffon (caracteristique du sexe femelle) par
rapport aux parapodes de la region posterieure transformee du porte-greffe.
forme atoque (lisse) alors que pendant le meme temps les cirres parapodiaux
dorsaux voisins et non amputes commencent a differencier des crenelures. Puis
les cirres parapodiaux dorsaux regeneres deviennent creneles avec l'hetero-
nereidation complete.
(b) Regeneration sur le greffon
Les tron?ons de corps fixes lateralement ou caudalement regenerent toujours
posterieurement et de facon synchrone par rapport au porte-greffe. De meme,
les parapodes et les cirres parapodiaux des greffons regenerent normalement
apres amputation (Figs. 3 a 6).
Les cirres parapodiaux regeneres, soit au niveau des segments neoformes par

La differentiation des
caracteres
sexuels
somatiques
chez Nereis 187
les troncons de corps fixes lateralement (97) ou caudalement (21) sur le porte-
greffe, soit apres ablation elective de cirres parapodiaux sur les greffons (127
troncons de corps; 136 bandes de parapodes) presentent dans tous les cas
{Nereis differenciees sexuellement ou non au moment de la greffe) les memes
caracteres sexuels que les cirres parapodiaux anciens du greffon.
(2) Parapodes surnumeraires
Nous considererons d'abord les conditions d'apparition de ces parapodes puis
leur comportement au cours de l'heteronereidation.
(a) Conditions d'apparition
des
parapodes surnumeraires
Des parapodes surnumeraires se sont edifies auniveau de la jonction greffon-
porte-greffe dans un certain nombre de cas de greffes de troncon de corps en
position laterale (a) et pour presque toutes les greffes de parapodes (/?).
(a) Cas de greffes laterales de trongon de corps
Greffe sur la face dorsale du porte-greffe. Des parapodes surnumeraires se
forment dans 45 % des cas environ (54/122) pour les troncons fixes par leur
partie anterieure ou par leur partie posterieure (8/19) sur la face dorsale du
porte-greffe. Le nombre de parapodes neoformes au niveau de la jonction
greffon-porte-greffe varie entre 1 a
5
parapodes. Neanmoins, tous les parapodes
se forment au niveau de la jonction de la face dorsale du porte-greffe et de la
face ventrale du greffon a la base des parapodes (70 % des parapodes surnume-
raires) ou au niveau de l'extremite de la chaine nerveuse sectionnee du greffon
jouxtant le porte-greffe (30 % des parapodes surnumeraires). Us sont orientes le
cirre ventral dirige vers la face ventrale du greffon et le cirre dorsal vers la face
dorsale du porte-greffe (Fig. 7).
Greffe sur la face ventrale
du
porte-greffe. Pour les greffes realisees sur la face
ventrale la neoformation de parapodes se produit dans tous les cas (9/9) au
niveau de la jonction des deux faces heterologues (dorsale du greffon, ventrale du
porte-greffe) a proximite des nerfs parapodiaux sectionnes du porte-greffe. Le
cirre ventral du parapode surnumeraire est alors oriente vers la face ventrale du
porte-greffe et le cirre dorsal vers la face dorsale du greffon.
(/?) Cas de greffes de parapodes
Les parapodes surnumeraires sont apparus dans tous les cas de greffes de
parapodes dans la region posterieure du corps (101 survivants) et dans 67%
des cas de greffes de parapodes dans la region anterieure (35 survivants).
L'etude de tous les cas de neoformations obtenues montre que:
- les parapodes neoformes sont generalement plus petits que les parapodes
anciens mais parfaitement constitues;
- pour un meme type de greffe, les parapodes apparaissent toujours sur le
meme cote du greffon et au niveau de contact entre greffon et porte-greffe;
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%