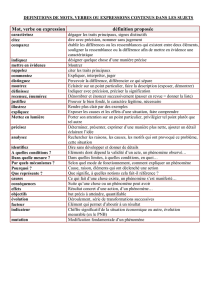
Mécanismes de variation des traits fonctionnels dans les prairies

M´ecanismes de variation des traits fonctionnels dans les
prairies des Alpes
Fabrice Grassein
To cite this version:
Fabrice Grassein. M´ecanismes de variation des traits fonctionnels dans les prairies des Alpes.
´
Ecologie, Environnement. Universit´e Joseph-Fourier - Grenoble I, 2009. Fran¸cais. <tel-
00560097>
HAL Id: tel-00560097
https://tel.archives-ouvertes.fr/tel-00560097
Submitted on 27 Jan 2011
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.

Université Joseph Fourier – Grenoble 1
Ecole Doctorale Chimie et Sciences du Vivant
THESE
Pour l‟obtention du titre de
Docteur en Biologie
Mention Biodiversité-Ecologie-Environnement
Mécanismes de variation des traits fonctionnels dans
les prairies des Alpes.
Par Fabrice Grassein
Soutenue le 23 octobre 2009
Membres du jury :
Marie Laure Navas – Professeur, ENSAM, Montpellier – Rapporteur
Laurent Hazard – Chargé de recherche, INRA, Toulouse – Rapporteur
Philippe Cozic – Directeur de recherche, CEMAGREF, Grenoble – Examinateur
Bill Shipley – Professeur, Université de Sherbrooke (Qc), Canada – Examinateur
Sandra Lavorel – Directeur de recherche, CNRS, Grenoble – Directrice de thèse
Irène Till – Directeur de recherche, CNRS, Grenoble – Directrice de thèse
Thèse réalisé au sein du Laboratoire d’Ecologie Alpine1 et de la Station Alpine Joseph
Fourier2
1 : UMR UJF-CNRS 5553 BP53 Université Joseph Fourier 38041 Grenoble Cedex 9
2 : UMS UJF-CNRS 2925 BP53 Université Joseph Fourier 38041 Grenoble Cedex 9


Avant-propos
Ce manuscrit déposé à l‟école doctorale Chimie et Sciences du vivant de l‟université de
Grenoble (Université Joseph Fourier) se présente sous la forme d‟une thèse sur articles. Il
comporte une introduction en français du contexte général et des méthodes employées, trois
chapitres regroupant les articles en anglais et une présentation succincte des objectifs et
résultats obtenus, et enfin une discussion sous la forme de synthèse des résultats obtenus.
Mon allocation de recherche pour trois ans a été financée par le Cluster de recherche
Rhône-Alpes Environnement. Ma quatrième année a été financée grâce à un poste d‟attaché
temporaire à l‟enseignement et à la recherche à l‟UFR de Biologie de l‟UJF. Le financement
des expérimentations a été rendu possible grâce au soutien financier du Cluster de recherche
Rhône-Alpes Environnement ainsi que par le biais du projet ANR-05-BDIV-009-01 QDIV
auquel le Laboratoire d‟Ecologie Alpine participait. Ce travail a été réalisé au sein du
Laboratoire d‟Ecologie Alpine de l‟université de Grenoble ainsi que sur les zones
expérimentales de la Station Alpine Joseph Fourier à Grenoble et au Col du Lautaret (Hautes
Alpes).

Avant-propos
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
1
/
245
100%