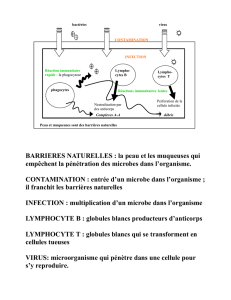
Activation et régulation des lymphocytes B : rôle de la costimulation

Le Courrier de la Transplantation - Vol. XIV - n° 1 - janvier-février-mars 2014
16
Dossier thématique
Rejet humoral
après transplantation
d’organe solide
Résumé
Summary
»
Les lymphocytes B nécessitent, pour le développement d’une
réponse anticorps spécifi que d’un antigène (Ag), l’intervention,
dans les organes lymphoïdes secondaires, de partenaires tels
que les cellules dendritiques (qui rapportent l’Ag capturé)
et les lymphocytes T CD4 auxiliaires. Ces derniers sont plus
particulièrement une sous-population CD4 retrouvée dans les
centres germinatifs des follicules lymphoïdes et appelée cellules T
folliculaires auxiliaires (T follicular Helper cells [TFH]). À partir d’une
même spécifi cité pour l’Ag partagée entre les cellules B et les
TFH, des interactions moléculaires de costimulation vont aussi
s’exercer pour parvenir à une activation complète du lymphocyte B :
CD80/86-CD28, CD40-CD40L, ICOS-ICOSL, PD1-PDL1/PDL2,
associées à d’autres facteurs tels que BAFF, IL-21 et IL-4. À l’issue
de cette activation primaire, demeurera une mémoire humorale.
Mots-clés : Lymphocyte B - Lymphocyte T auxiliaire (TFH) –
Costimulation.
In order to develop an antigen (Ag) specific antibody
response, B lymphocytes need the intervention of other
partners such as dendritic cells (that capture the Ag)
and CD4 helper T lymphocytes. These CD4 cells are more
specifically a sub-population located in the germinal centers
of the lymphoid follicles, subsequently called T Follicular
Helper cells (TFH). From an identical Ag specificity, shared
by B and TFH cells, costimulation molecular interactions
will take place, such as CD80/86-CD28, CD40-CD40L, ICOS-
ICOSL, PD1-PDL1/PDL2 , in addition to other factors such
as BAFF, IL-21 and IL-4. Following this primary activation
will remain a humoral memory.
Keywords: B lymphocyte - Helper T lymphocyte –
Costimulation.
Activation et régulation des lymphocytes B :
rôle de la costimulation
Activation and regulation of
B
lymphocytes: role of the costimulation
Gilles Blancho*
Les lymphocytes B sont impliqués dans diverses
fonctions, au premier rang desquelles se situe
la synthèse des anticorps (Ac) ou des immuno-
globulines (Ig). Cette synthèse répond à la stimulation
par 2 types d’antigènes (Ag), les Ag T indépendants
et T dépendants. Les T indépendants, ainsi que leur
appellation l’indique, ne nécessitent pas l’intervention
de lymphocytes T pour l’induction des Ig, correspondent
plutôt à des mitogènes de type polymères de séquences
répétées tels que le LPS (lipopolysaccharide) ou le
pokeweed mitogen (PWM) et induisent essentiellement
une activation lymphocytaire B précoce non spécifi que
d’un Ag, voire spécifi que mais de faible affi nité. Les
autres types d’Ag, en général de nature glycoprotidique,
vont induire une réponse qui leur est spécifi que, en
faisant intervenir une aide intermédiaire, le lymphocyte
T CD4 dit auxiliaire (T helper). Cette aide va se produire
essentiellement dans les zones lymphoïdes secondaires
(ganglions lymphatiques et rate) et à travers des inter-
actions moléculaires intercellulaires dans le contexte
d’une synapse dite immunologique. Après la rencontre
de l’Ag, certaines interactions seront nécessaires pour
aller jusqu’à une vraie activation ; on parle d’interactions
de costimulation.
La réponse humorale
dans les zones lymphoïdes
Le terme “humoral” qualifie des composants non
cellulaires des systèmes circulatoires sanguin et lym-
phatique et, de fait, en matière de réponse immune,
se rapporte donc aux Ig. Ces dernières sont capables
de reconnaître des Ag circulants de façon conforma-
tionnelle (non apprêtés) mais spécifi quement, car leur
cellule primordiale, le lymphocyte B ou son expression
diff érenciée ultime, le plasmocyte, ont été recrutés
* Institut
detransplantation-urologie-
néphrologie (ITUN),
CHU de Nantes.
CT-n1-janv-fév-mars 2014-B.indd 16 26/03/14 11:35

Le Courrier de la Transplantation - Vol. XIV - n° 1 - janvier-février-mars 2014 17
dans leur fonction à synthétiser des Ig de façon spé-
cifi que. Ce recrutement se déroule donc en amont à
partir d’un lymphocyte B naïf et met à contribution les
cellules dendritiques et les lymphocytes T. D’un côté,
dans les zones lymphoïdes T, les cellules dendritiques
rapportent et présentent l’Ag, en particulier par leurs
molécules du complexe majeur d’histocompatibilité
(CMH) de classe II, aux lymphocytes CD4 porteurs d’un
récepteur T (TCR) capable de reconnaître cet Ag. D’un
autre côté, dans les zones lymphoïdes B, les macro-
phages des sinus sous-capsulaires captent les Ag et les
présentent aux lymphocytes B qui vont les capter par
leurs récepteurs à l’Ag (B Cell Receptor [BCR]) qui sont
en fait des Ig de membrane. Ces dernières vont ainsi
internaliser l’Ag pour, après apprêtement, le présenter
par leurs molécules du CMH de classe II aux lympho-
cytes CD4 préalablement recrutés précisément pour
leur spécifi cité à ce même Ag dans les zones intermé-
diaires entre les zones T et B et fournissant alors cette
aide à ces lymphocytes B. Cette coopération CD4 sera
un deuxième signal donné à la cellule B, à la suite du
premier signal d’engagement du BCR, qui consistera
en des interactions moléculaires et des libérations de
certaines cytokines. Le premier signal BCR aura cepen-
dant été indispensable, bien sûr pour engager la spé-
cifi cité de la réponse mais aussi pour transformer le
lymphocyte B en une véritable cellule présentatrice
de l’Ag (CPA), en particulier par une surrégulation de
l’expression des molécules CD80 et CD86 interagis-
sant avec la molécule CD28 du lymphocyte T. Il s’agit
là d’un des signaux de costimulation les plus précoces
et essentiels à la coopération T-B et donc à l’induction
d’une réponse humorale primaire.
Il apparaît maintenant que les lymphocytes CD4,
jouant ce rôle de coopération, à partir du précurseur
commun dans un environnement cytokinique parti-
culier, se diff é rencient en diverses sous-populations
appelées Th, numérotées selon leur expression de
facteurs transcriptionnels et de cytokines et exerçant
des fonctions auxiliaires spécialisées. Ainsi, dérivées
dans un environnement IL-12, les Th1 expriment T-bet,
Stat1 et Stat4 et sécrètent IFNγ, IL-2 et TNFα. Dans
un environnement IL-4, les CD4 se diff érencient en
Th2, exprimant GATA3, Stat5 et Stat6 et sécrétant IL-4,
IL-5, IL-13 et IL-10. Dans un environnement TGFβ et
IL-4, des Th9 vont se diff érencier en exprimant PU.1
(Spi-1) et en sécrétant IL-9 et IL-10. Dans un contexte
majoritairement IL-21, des Th17 vont se diff érencier
en exprimant RORγt, RORα et Stat3 et en sécrétant
plus particulièrement IL-17, IL-21 et IL-22. Enfi n, dans
la région du follicule lymphoïde et en particulier du
centre germinatif (CG) et sous infl uence IL-21, IL-6, des
lymphocytes dits “folliculaires auxiliaires” (T Follicular
Helper [TFH]) vont se diff érencier. Il apparaît que cette
sous-population, décrite plus récemment, est la plus
spécialisée dans la coopération avec le lymphocyte B
dans les follicules lymphoïdes, aboutissant essentiel-
lement à la formation et au maintien des CG et donc
à l’affi nement de la réponse Ac et à la diff érenciation
en plasmocytes et cellules mémoires.
Les lymphocytes T folliculaires auxiliaires,
ou Helper (TFH)
À la faveur d’une plus grande caractérisation des sous-
populations lymphocytaires, il est apparu que ces
lymphocytes CD4, coopérant avec les lymphocytes B
dans les CG des follicules lymphoïdes, exprimaient beau-
coup plus fortement la molécule CXCR5, répondant
ainsi à la chémoattraction de CXCL13 (tout comme
les cellules B pour les focaliser dans les follicules), per-
mettant une colocalisation T-B essentielle à la coopé-
ration T-B. Par la suite, il a été montré que ces cellules,
par rapport aux non-TFH, présentaient aussi une forte
expression du facteur de régulation transcriptionnelle
Bcl6, de PD-1, de SAP (SLAM-Associated Protein), d’IL-21
et d’ICOS (Inducible T Cell Costimulator) et, en revanche,
une absence d’expression de Blimp-1. Ainsi se caracté-
rise donc la sous-population TFH, si importante dans
la coopération mais aussi dans le maintien des CG des
follicules lymphoïdes et la régulation de la diff éren-
ciation B en plasmocytes et cellules B mémoires. Il se
trouve que Bcl6 et Blimp-1 sont des facteurs d’inhi-
bition mutuelle, à tel point que les lymphocytes CD4
exprimant Bcl6 (réprimant Blimp-1) sont des TFH, alors
que les CD4 exprimant Blimp-1 (réprimant Bcl6) sont
des non-TFH (1).
Il est par ailleurs probable que les TFH aient d’autres
fonctions que la coopération B directe, en interagissant
aussi avec la cellule dendritique folliculaire résidente
des follicules lymphoïdes (comme son nom l’indique)
et avec le troisième type cellulaire majeur des CG.
Une fois le CG formé, les TFH seront nécessaires pour
envoyer les signaux de survie et de prolifération des
cellules B du CG et stimuler l’hypermutation soma-
tique, véritable processus de maturation d’affi nité
des Ac générés dans le CG. Les diff érents signaux de
survie donnés par la TFH passent par diverses voies
comprenant CD40, IL-4, IL-21, PD-1, BAFF, entrant
en compétition avec l’interaction Fas-FasL induisant
l’apoptose des lymphocytes B en leur absence. De par
sa colocalisation avec les cellules B dans le CG, la TFH
va exercer ces interactions directes, que nous allons
brièvement revoir.
Activation et régulation des lymphocytes B : rôle de la costimulation
CT-n1-janv-fév-mars 2014-B.indd 17 26/03/14 11:35

Le Courrier de la Transplantation - Vol. XIV - n° 1 - janvier-février-mars 2014
18
Dossier thématique
Rejet humoral
après transplantation
d’organe solide
costimulation, mais plutôt un facteur de survie du
lymphocyte B interagissant sur son récepteur BAFF-R.
Avec le signal BCR, BAFF est un élément essentiel de
l’homéostasie des lymphocytes B. Avec le signal BCR,
BAFF est un élément essentiel de l’homéostasie des
lymphocytes B en association avec une autre molécule,
APRIL. Ces 2 molécules, essentiellement synthétisées
par des cellules de l’immunité innée, fi xent 3 types de
récepteurs (TACI, BCMA et BAFF-R) avec des affi nités
variables, que, afi n de rester dans le domaine de la
costimulation et non pas de la survie, nous ne déve-
lopperons pas ici.
IL-21
Cette cytokine est essentielle à la survie et la prolifé-
ration des cellules B des CG en agissant directement
sur leur récepteur à IL-21. Il s’agit en fait d’une action
duale, puisque la survie des TFH nécessite la présence
des B qui, eux, nécessitent IL-21. Son action complexe
passerait notamment par une surrégulation de Bcl-6
dans les B des CG. De plus, il s’agit de la cytokine dont
la fonction de diff érenciation plasmocytaire est la plus
puissante.
IL-4
Depuis longtemps, IL-4 était considérée comme un
facteur de survie et de diff érenciation du lymphocyte B,
notamment dans le contexte de la description initiale
de la sous-population Th, en tant que cytokine majeure
des Th2. Aujourd’hui, il apparaît clairement que ce
sont les TFH et non les Th2 qui jouent ce rôle, en étant
elles-mêmes capables de la sécréter. IL-4 joue un rôle
antiapoptotique crucial pour les cellules B du CG en
induisant une expression de Bcl-2.
Bien entendu, le schéma n’est pas aussi exclusif dans
les CG ; on retrouve en eff et l’intervention d’autres
cytokines telles que IFNγ, si important pour la diff éren-
ciation Th1. Précisément, des lymphocytes Th1 et Th2
peuvent aussi être retrouvés au milieu des comparti-
ments TFH. En outre, toute cette activité complexe des
CG est aussi régulée pour prévenir tout emballement
de la prolifération B. Ainsi, des cellules T régulatrices
Foxp3+ d’origine thymique ont été mises en évidence
dans les CG au point d’être défi nies comme Tfr. Elles
partagent cependant, la même expression en CXCR5,
PD1, ICOS et Bcl6, se colocalisent aussi dans les CG mais
expriment Foxp3, CTLA-4, GITR et IL-10 sans exprimer
les facteurs de coopération des cellules B, IL-21, IL-4
et CD40L. Ainsi présentent-elles des fonctions sup-
pressives aussi bien sur le niveau des réponses dans
les CG, la prolifération des B dans les CG que la pro-
duction d’Ac (3).
Les interactions TFH-lymphocyte B
dans le centre germinatif
CD80/86-CD28
Cette interaction bien connue comme interaction
de costimulation principale ou second signal entre
les CPA, en particulier les cellules dendritiques, et les
lymphocytes T (2), s’avère tout aussi importante dans les
réponses en aval : sans une ligation CD28, la cellule T ne
reçoit pas de signal d’activation et il ne pourra y avoir
de génération de TFH.
CD40L-CD40
CD40 est exprimé par le lymphocyte B et interagit avec
son ligand spécifi que, CD40L, fortement exprimé par
les T CD4. Cette interaction joue un rôle majeur, tant
son absence empêche le développement des CG et
des plasmocytes. L’interaction globale va plutôt faire
entrer le lymphocyte B en prolifération et, dans une
zone de très fort niveau d’apoptose, en épargner ce
dernier probablement par une surrégulation de Bcl6.
En retour, le lymphocyte T, via le signal CD40L, va libérer
certaines cytokines, en particulier IL-21, IL-4 et proba-
blement aussi IFNγ, jouant des rôles importants dans
l’orientation de la commutation de classe (changement
d’isotype des Ig) et le maintien des CG. La nécessité
d’une interaction physique CD40-CD40L locale garantit
une spécifi cité de la réponse B, puisqu’elle se produira
seulement entre cellules spécifi ques de l’Ag.
ICOS-ICOSL
Le signal ICOS est absolument nécessaire pour une
génération optimale des TFH, mais pas des autres sous-
populations CD4. Après le signal CD28, la molécule ICOS
va se surréguler sur bon nombre des CD4 mais surtout
sur les TFH. La genèse et le maintien de ces derniers
vont ensuite nécessiter impérativement l’interaction
avec ICOSL pour leurs genèse et maintien, vraisem-
blablement par l’induction de l’expression de Bcl-6.
PD1-PD-L1/PD-L2
PD1 (Programmed Death 1) est un récepteur inhibiteur
puissant de la TFH. Il interagit avec PD-L1 et PD-L2,
exprimés par les cellules B des CG. Il semble que ce
haut niveau d’expression par les TFH soit entretenu par
les interactions T-B dans le CG et que ses eff ets soient
doubles, avec un signal d’activation aux lymphocytes
B des CG mais aussi d’interruption des interactions T-B
par inhibition du signal stop du TCR.
BAFF
BAFF (B cell Activating Factor belonging to the TNF
Family) n’est pas à proprement parler un facteur de
CT-n1-janv-fév-mars 2014-B.indd 18 26/03/14 11:35

Le Courrier de la Transplantation - Vol. XIV - n° 1 - janvier-février-mars 2014 19
Induction de la mémoire humorale
Une des caractéristiques de cette organisation de la
réponse B, en particulier dans les CG, est d’induire une
mémoire fondée sur les Ac, à partir de la génération
de cellules B mémoires à longue vie. Leur activation
nécessite toujours l’intervention de cellules T auxi-
liaires, elles-mêmes de phénotype mémoire, les TFH
mémoires. À la suite d’une restimulation antigénique,
les B mémoires vont représenter l’Ag spécifi que aux TFH
mémoires qui vont promouvoir leur expansion clonale,
leur diff érenciation rapide en plasmocytes mémoires et
l’induction de CG secondaires. Il s’agit toujours d’inte-
ractions moléculaires locales, tant les TFH mémoires
expriment toujours CXCR5, qui les maintient dans les
CG. Il semble que les B mémoires, porteurs de BCR de
haute affi nité pour l’Ag, vont être capables de capter de
très faibles quantités d’Ag et de les présenter comme de
véritables CPA aux TFH mémoires qui, en retour, vont
surréguler leurs signaux cytokiniques (plutôt au repos
en absence de l’Ag) avec les conséquences sus-citées (4).
Conclusion
La costimulation B implique des interactions essentielles
T-B dans les CG des follicules lymphoïdes. De véritables
synapses immunologiques s’y forment, assurant la spé-
cifi cité de la réponse antigénique et mettant en jeu des
interactions moléculaires de costimulation, d’adhésion
et de suppression/régulation, permettant d’activer puis
moduler la réponse B, tout en initiant la mise en place de
sa mémoire, si importante dans l’immunocompétence
d’un individu.
■
1.
Crotty S. Follicular Helper CD4 T cells (TFH).
Annu Rev Immunol 2011;29:621-63.
2. Poirier N, Blancho G, Vanhove B. CD28-
specifi c immunomodulating antibodies: what
can be learned from experimental models?
Am J Transplant 2012;12(7):1682-90.
3. Qi H. From SAP-less T cells to helpless B
cells and back: dynamic T-B cell interactions
underlie germinal center development and
function. Immunol Rev 2012;247:24-35.
4.
McHeyzer-Williams M, Okitsu S, Wang N,
McHeyzer-Williams L. Molecular program-
ming of B cell memory. Nat Rev Immunol
2011;12(1):24-34.
Références bibliographiques
L’auteur déclare ne pas avoir de
liens d’intérêts.
SFT
Le 13e congrès annuel de la SFT
à Marrakech (11-14 décembre 2013)
a rassemblé 571 participants, dont 179 paramédicaux
(DIU, journée paramédicale et École francophone de prélèvement multi-organe).
Activation et régulation des lymphocytes B : rôle de la costimulation
CT-n1-janv-fév-mars 2014-B.indd 19 26/03/14 11:36
1
/
4
100%