cours 13 : le cortex sensoriel et la plasticité du

COURS 13 : LE CORTEX SENSORIEL ET LA PLASTICITÉ DU
SYSTÈME NERVEUX CENTRAL
Le chapitre précédent a mis en évidence l'organisation fonctionnelle des circuits neuroniques assurant la
réalisation d'un phénotype comportemental : le réflexe myotatique.
Dans ce chapitre, on cherche à mettre en évidence les parts respectives du génotype et de
l’environnement dans la réalisation des phénotypes comportementaux.
Malgré la mise en place d’un plan d’organisation du système nerveux commun à tous les vertébrés et le
contrôle génétique du développement du réseau neuronique, chaque individu ne perçoit pas son
environnement de la même façon.
I) La part du génotype dans le fonctionnement du système nerveux
A) La part du génotype dans la mise en place des réseaux de neurones
On connaît chez la Souris un certain nombre de mutations qui touchent le système nerveux. Un groupe de
mutants montre des anomalies du développement post-natal d'une région de l’encéphale : le cervelet. De
tels mutants sont appelés mutants cérébelleux.
Le cervelet est une région de l'encéphale qui joue un rôle dans la coordination des mouvements.
Divers noms sont donnés à ces mutants : tournoyant, chancelant, titubant, selon les altérations
locomotrices qui les caractérisent. Tous ces mutants présentent, à l’échelle du phénotype cellulaire, une
désorganisation des circuits de neurones dans le cortex de leur cervelet.
Pour chaque type de mutant, l'anomalie est due à une mutation d’un des gènes codant pour des protéines
impliquées dans la mise en place des réseaux de neurones.
Au niveau du SNC, les aires de projection varient d’une espèce à l’autre (homonculi somatosensoriels
différents chez l’Homme, le rat, le singe...) mais au sein d’une même espèce les aires sensorielles et
motrices sont localisées aux mêmes endroits, suggérant l’intervention du génotype. La répartition en
couches et colonnes du cortex cérébral et l’apparition de mutants qui présentent les couches I à VI
dans le désordre prouvent l’intervention du génotype dans l’établissement des phénotypes micro et
macroscopique du cortex cérébral. D’autre part, la dominance hémisphérique gauche pour le
langage et droite pour la perception visuo-spatiale serait également génétiquement déterminée. Le
cerveau gauche serait plus analytique et le cerveau droit plus globaliste dans son fonctionnement.
L’étude du daltonisme, de l’insensibilité gustative et de maladies rares comme certaines formes
d’insensibilité́ congénitale à la douleur, dont l’origine est une mutation génétique, montre que le système
nerveux périphérique (SNP) est également, en partie, sous contrôle génétique.
Des mutations des gènes homéotiques Hox-1.3 et Pax-3 entraînent des malformations des SNC et SNP.
B) La part du génotype dans la réalisation d’un réflexe
On connaît des cas d'insensibilité congénitale à la douleur, sans altération des autres fonctions
sensorielles.
L'insensibilité congénitale à la douleur est une maladie exceptionnelle, le plus souvent de cause
génétique, caractérisée par une incapacité à percevoir la douleur sous toutes ses formes et sur tout le
corps, avec conservation des autres sensations tactiles.
Certaines formes de la maladie sont dues à une augmentation de la production d'endorphines dans le
cerveau.
Une mutation du gène codant pour le récepteur au NGF (le NGF = Nerve Growth Factor, est un facteur de
croissance indispensable au développement des neurones) est responsable de cette grave maladie. Chez
les individus possédant cette mutation, les fibres amyéliniques (dépourvues de gaine de myéline)

normalement responsables de la conduction des messages douloureux ne se mettent pas en place lors du
développement, ce qui rend impossible la réalisation du réflexe nociceptif (le stimulus est la douleur).
Des exemples de mutations perturbant l'organisation des réseaux neuroniques sont connus et démontrent
la part du génotype dans la réalisation d'un phénotype comportemental :
• Certaines mutations ont pour effet de modifier la façon dont les neurones impliqués dans le contrôle de la
motricité sont interconnectés dans l'encéphale : la mise en place des réseaux de neurones est donc
sous contrôle du génotype.
• Une mutation peut empêcher la croissance de fibres sensorielles qui, normalement assurent la
transmission de messages nerveux impliqués dans des réflexes : le développement de ces fibres est
donc sous contrôle du génotype.
Un phénotype comportemental est donc la conséquence de la mise en place, au cours du développement,
des chaînes de neurones, sous le contrôle de l'information génétique.
Les phénotypes microscopiques et macroscopiques des homonculi, de la répartition en couches et colonne
du cortex cérébral (au niveau du SNC), le phénotype comportemental des réflexes (reflexes myotatique,
nociceptif) comme la sensibilité́ de chacun à certains goûts, certaines couleurs, a la douleur (au niveau du
SNP) sont la conséquence directe de la mise en place, au cours du développement embryonnaire, des
chaînes neuroniques, sous le contrôle du génotype.
Cette mise en place des réseaux de neurones s’effectue généralement sous l’action d’hormones comme le
NGF (Nerve Growth Factor) qui orientent la croissance des neurones.
II) Expérience individuelle et plasticité du système nerveux
L'étude du cortex cérébral permet de montrer l’influence de l'environnement sur le fonctionnement du
système neveux. On appelle cortex cérébral la partie superficielle des hémisphères cérébraux, formée par
de la substance grise. Le terme « cerveau» au sens strict désigne les hémisphères cérébraux.

A) La somesthésie
1) Définition
La somesthésie représente l'ensemble des mécanismes nerveux recueillant l’information
sensorielle du corps.
La somesthésie désigne donc un ensemble de différentes sensations (pression, chaleur, douleur ...) qui
proviennent de plusieurs régions du corps (peau, tendons, articulations, viscères …). Ces sensations sont
élaborées à partir des informations fournies par de nombreux récepteurs sensitifs du système
somatosensoriel (ou somesthésique), situés dans les tissus de l'organisme.
2) Classification des modalités somesthésiques
Selon la nature du stimulus, on distingue trois modalités somesthésiques principales :
-la sensibilité mécanoréceptrice, mise en jeu par toute stimulation mécanique de l'organisme (ou
proprioception), (proprioception = qui se rapporte à la sensibilité du système nerveux aux informations
provenant des muscles, des articulations et des os).
-la sensibilité douloureuse ou nociception, mise en jeu par tout stimulus provoquant une lésion tissulaire,
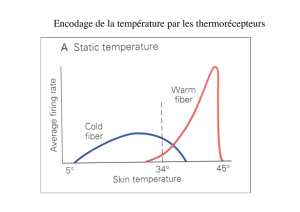
-la sensibilité thermique, répondant au chaud et au froid.
3) Les mécanorécepteurs
Ils sont à l’origine de toutes les sensations tactiles et de la sensibilité proprioceptive qui contribue à
déterminer la position de notre corps dans l'espace, de nos membres par rapport au corps : sensibilité à la
position, au mouvement (kinesthésie) et à la force.
Les récepteurs tactiles :
Il en existe différents types morphologiques, répartis dans le derme et l'épiderme et responsables de
différentes sensibilités tactiles à la surface du corps : pression, chatouillement, vibration. On distingue :
- les terminaisons nerveuses libres
- les corpuscules de Meissner
- les disques de Merkel,
- les récepteurs pileux,
- les corpuscules de Ruffini et de Pacini.

Terminaisons nerveuses libres : présentes pratiquement partout dans la peau, fibres non myélinisées.
Les corpuscules de Meissner : formés d’une fibre sensorielle myélinisée de gros calibre dont la
terminaison très ramifiée est enfermée dans une capsule. On les trouve dans la peau glabre, les lèvres, le
bout des doigts et autres zones de la peau à haute sensibilité tactile.
Les disques de Merkel : on les trouve souvent dans les mêmes régions que les corpuscules de Meissner,
et dans la peau pileuse. Ce sont des cellules épithéliales spécialisées connectées à une fibre nerveuse.
Les récepteurs pileux : fibre enroulée à la base du poil.
Les corpuscules de Ruffini : terminaison multiramifiée sensible à la déformation ; on les trouve dans les
couches les plus profondes de la peau.
Les corpuscules de Pacini : sensibles aux vibrations, on les trouve également dans les couches les plus
profondes de la peau (hypoderme).
Les récepteurs des muscles, des tendons et des articulations :
Dans les muscles squelettiques, on trouve les fuseaux neuromusculaires (se compose de 3 à 10 petites
fibres musculaires modifiées, les fibres intrafusales, reliées à une terminaison dendritique), dans les
tendons, les organes tendineux de Golgi.
Ce sont des récepteurs sensoriels spécialisés, sensibles aux variations de longueur du muscle ou du
tendon. On trouve dans les articulations des corpuscules de Ruffini sensibles aux déformations et qui
détectent la position de l’articulation.

Le FNM
Les propriocepteurs
4) Les récepteurs à la douleur
On les appelle aussi nocicepteurs. Ce sont des terminaisons nerveuses libres, présentes dans les
couches superficielles de la peau et dans certains tissus profonds comme les méninges, les parois
artérielles, etc.
Ils sont sensibles à des stimulations de nature physique variée : mécaniques, chimiques, thermiques.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
6
7
8
9
10
11
12
13
14
15
16
17
18
19
1
/
19
100%