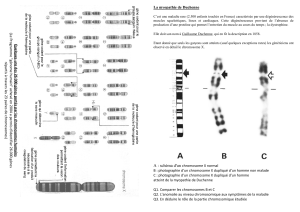
L`avenir du chromosome Y

Journal Identification = MTE Article Identification = 0442 Date: March 15, 2013 Time: 4:9 pm
Mini-revue
mt Médecine de la Reproduction, Gynécologie Endocrinologie 2013 ; 15 (1) : 78-86
L’avenir du chromosome Y
Human sex chromosomes evolution: is the Y disappearing?
Émilie Lecompte1,2
1Université de Toulouse,
UMR5174 EDB,
laboratoire évolution et diversité
biologique,
118, route de Narbonne,
31062 Toulouse,
France
2CNRS,
université Paul-Sabatier,
UMR5174 EDB,
31062 Toulouse,
France
Résumé. Les comparaisons de génomes entre vertébrés démontrent que les chromosomes
sexuels humains dérivent d’un système mis en place chez les mammifères il y a quelque
150 millions d’années avec un déterminisme du sexe mâle, spécifique, dépendant du gène
Sry. La différenciation X-Y et la spécialisation du Y dans la fonction mâle sont le résultat d’une
longue évolution à partir d’une paire d’autosomes, suite à la suppression de la recombinai-
son entre le X et le Y. L’arrêt de la recombinaison a également entraîné la perte de gènes,
l’accumulation de mutations et de séquences répétées conduisant à la dégénérescence du Y.
La dégradation va-t-elle se poursuivre inexorablement, jusqu’à ce que le Y disparaisse ? Les
arguments pour et contre cette disparition chez l’homme sont explicités. Des mécanismes
pouvant conduire à la disparition du Y sont illustrés grâce à l’étude d’espèces de mammifères
sans Y.
Mots clés : chromosome Y, dégénérescence, évolution, déterminisme du sexe
Abstract. Genome comparisons between vertebrates show that human sex chromosomes evol-
ved from a mammalian male sex-determining system that arose about 150 million years ago,
dependant of a specific gene, Sry. The X-Y differentiation and Y specialization in male sex
and fertility are the result of a long evolution from an autosomal pair, after the recombination
stops between the X and Y. The recombination suppression drives gene loss, accumulation of
mutations and repeated sequences, leading to the Y degeneration. Should Y degrade inexo-
rably, until its demise? Arguments for and against the human Y disappearance are explained.
Researches on the mammalian species without Y chromosome exemplify some mechanisms
allowing Y loss.
Key words: Y chromosome, degeneration, evolution, sex determination
Déterminisme du sexe
et chromosomes sexuels
humains
Depuis leur découverte, les chro-
mosomes sexuels ont été source
de fascination et l’objet de nom-
breuses recherches. Cette découverte
remonte aux débuts de la cytogé-
nétique lorsqu’en 1912, Winiwater
constata que la femme présentait un
chromosome X en double exemplaire
alors que celui-ci n’était présent qu’en
un exemplaire chez l’homme. En
1921, Painter découvrit le Y, passé
inaperc¸u jusqu’alors du fait de sa
petite taille, puis conclut en 1923 à
un mécanisme du déterminisme du
sexe lié à la formule XX ou XY. Dès
lors, de nombreuses études ont porté
tant sur le déterminisme du sexe que
sur les mécanismes d’évolution ayant
conduit à la paire XY.
La recherche médicale s’est sur-
tout intéressée à déchiffrer la cascade
génétique complexe de déterminisme
du sexe à l’origine du développement
de l’embryon, soit en mâle, soit en
femelle. L’étude de souris mutantes ou
de pathologies humaines, comme le
syndrome de Klinefelter (XXY) ou de
Turner (X0), a permis l’identification
de nouveaux gènes impliqués dans la
voie de différenciation des gonades et
mieux comprendre les relations entre
les protéines impliquées. Les avancées
les plus importantes dans ce champ de
recherche ont été l’identification du
gène de déterminisme du sexe mâle,
Sry [1]. Depuis, de nombreux gènes
liés au sexe ont été décrits, comme
Sox9 ayant un rôle déterminant dans
la différenciation en mâle [2, 3] ;
d’autres restent encore à découvrir.
On en sait finalement assez peu sur
le mode d’action et l’identité des
nombreux gènes impliqués dans la
cascade de déterminisme du sexe et
en aval de cette cascade [4].
De son côté, la biologie évolutive
s’est intéressée à la question fonda-
mentale de l’origine et de l’évolution
doi:10.1684/mte.2013.0442
médecine thérapeutique
Médecine
de la Reproduction
Gynécologie
Endocrinologie
Tirés à part : E. Lecompte
78 Pour citer cet article : Lecompte É. L’avenir du chromosome Y. mt Médecine de la Reproduction, Gynécologie Endocrinologie 2013 ; 15 (1) : 78-86
doi:10.1684/mte.2013.0442
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

Journal Identification = MTE Article Identification = 0442 Date: March 15, 2013 Time: 4:9 pm
des chromosomes sexuels et a ainsi apporté des éclaircis-
sements sur le déterminisme du sexe. Chez la majorité
des vertébrés, les mâles et les femelles sont séparés,
pourtant le sexe peut être déterminé par différents sys-
tèmes. Dans certaines espèces, le sexe n’est pas déterminé
génétiquement mais conditionné par l’environnement,
comme, par exemple, la température d’incubation des
œufs chez les crocodiles et les tortues marines. Par ailleurs,
il existe une grande diversité de systèmes différents parmi
les espèces possédant des chromosomes sexuels [5].
La plupart des gènes impliqués dans le déterminisme
du sexe sont conservés chez l’ensemble des vertébrés,
comme DMRT1 ou Sox9, tandis que Sry est spécifique
aux mammifères thériens (marsupiaux et placentaires)
[2, 6, 7].
Chez les oiseaux, ce sont les femelles qui sont hété-
rogamétiques, avec des chromosomes sexuels différents
nommés Z et W. Parmi les lézards, certaines espèces pré-
sentent le système XY, d’autres ZW et chez d’autres encore,
le sexe est déterminé par la température d’incubation [5].
À l’échelle de l’évolution des vertébrés, la comparaison de
génomes à large échelle a permis d’identifier les homolo-
gies entre les chromosomes. Par exemple, la comparaison
des génomes de l’homme et du poulet a démontré que
l’homologue de notre X chez les oiseaux est un auto-
some et non un chromosome sexuel ; réciproquement,
les chromosomes sexuels des oiseaux sont homologues à
des autosomes de l’homme [8]. Ainsi, il apparaît que les
chromosomes sexuels des mammifères (XY), des oiseaux
(ZW) ou des serpents (ZW) ne sont pas homologues et ont
donc évolué indépendamment [8, 9]. Les chromosomes
XY de l’homme sont homologues à ceux de l’ensemble des
mammifères thériens (placentaires et marsupiaux) alors
que les monotrèmes (ornithorynques et échidnés) pos-
sèdent cinq paires de chromosomes XY qui ne sont pas
homologues au X et Y humains [10]. Nos chromosomes
sexuels sont donc apparus après la divergence des mono-
trèmes, soit ilyaenviron 150 millions d’années (Ma) à
partir d’un système ancestral ZW [10, 11]. Cependant,
les chromosomes sexuels des marsupiaux sont bien plus
petits et la majeure partie du X et Y des placentaires est
homologue d’une région autosomale des marsupiaux. Ces
homologies permettent de définir deux régions, une région
ancienne, conservée chez tous les mammifères thériens, et
une région ajoutée plus récemment sur le X et le Y des seuls
placentaires et qui regroupe la plupart des gènes actuels
du Y humain [12].
Ainsi, l’identification des chromosomes homologues
parmi les vertébrés montre que les chromosomes sexuels
humains dérivent d’un système mis en place chez les
mammifères thériens avec un déterminisme du sexe mâle,
spécifique, dépendant du gène Sry. La différenciation du X
et du Y et la spécialisation du Y dans la fonction mâle sont
le résultat d’une longue évolution et de divers mécanismes
qui vont maintenant être explicitées.
Différenciation et spécialisation
du chromosome Y humain
Les chromosomes X et Y sont très différents : le X est
un grand chromosome (155 Mb chez l’homme) portant
1 098 gènes alors que le Y est petit (64 Mb chez l’homme)
et ne contient que 106 gènes [13, 14]. De plus, le Y
humain est très hétérogène et comporte différents types de
séquences, situées sur deux régions distinctes d’un point
de vue évolutif : les régions pseudo-autosomales (RPA) qui
comprennent 28 gènes du Y (∼26 %) et la région non
recombinante, appelée région mâle-spécifique du chro-
mosome Y (MSY) [13]. Les RPA constituent de petites
régions situées à l’extrémité du Y permettant l’appariement
de la paire, lors de la méiose. La MSY, largement hété-
rochromatique (∼40 Mb), transmise de fac¸on clonale de
père en fils, est spécialisée dans la fonction mâle (déter-
minisme du sexe et spermatogenèse).
Les différences importantes entre les contenus
géniques des chromosomes sexuels peuvent affecter le
dosage de nombreux gènes entre les sexes et pro-
duire des conséquences phénotypiques sérieuses. Divers
mécanismes de régulation ont été découverts chez les
lignées présentant des chromosomes sexuels et des dif-
férences de dosage entre les mâles et les femelles [15].
Chez les mammifères, la compensation de dosage se
réalise par l’inactivation d’un chromosome X chez les
femelles.
Bien que les X et Y humains soient aujourd’hui très
différents par leur taille et leur contenu génique, ils ont
une origine unique et dérivent d’une paire d’autosomes
qui aurait suivi une évolution complexe. La divergence
entre le X et le Y a commencé après l’acquisition, sur un
chromosome de la paire, le proto-Y, d’un gène de déter-
minisme du sexe mâle, Sry, qui a évolué à partir de son
homologue Sox3 présent sur le X [16]. La suppression de
la recombinaison est considérée comme l’étape cruciale
de l’évolution des chromosomes sexuels puisqu’elle per-
met l’accumulation de mutations spécifiques à un seul
chromosome, favorisant l’émergence de facteurs sexe-
spécifiques [17]. La spécialisation du Y dans la fonction
mâle semble avoir été réalisée par l’évolution de quelques
gènes initialement présents sur le Y vers un rôle dans
la fonction mâle [18]. Le maintien d’une combinaison
de gènes et/ou d’allèles critiques pour le déterminisme
du sexe et l’antagonisme sexuel n’est en effet possible
qu’en absence de recombinaison. En effet, les mutations
avantageuses pour les mâles sont désavantageuses pour
les femelles et sont généralement contre-sélectionnées
puisqu’elles diminuent les chances de reproduction des
femelles (antagonisme sexuel). L’accumulation de fonc-
tions spécifiques des mâles sur le Y aurait ainsi favorisé
l’extension des zones non recombinantes réduisant la
région recombinante aux extrémités des chromosomes X
et Y (les RPA), probablement du fait de l’accumulation
mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 15, n◦1, janvier-février-mars 2013 79
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

Journal Identification = MTE Article Identification = 0442 Date: March 15, 2013 Time: 4:9 pm
Mini-revue
de remaniements chromosomiques tels que des inver-
sions [19, 20]. Cette hypothèse a été validée notamment
grâce aux comparaisons de génomes et au séquenc¸age
des chromosomes X et Y humains [13, 14]. La portion
euchromatique de la MSY humaine, relativement petite
(∼23 Mb), contient 172 unités transcriptionnelles codant
pour 27 protéines différentes, la plupart étant exprimées
uniquement dans les testicules et impliquées dans la sper-
matogenèse [13, 21].
Les séquences localisées dans la MSY révèlent des his-
toires évolutives très différentes, tant au regard de leur
origine que de leur évolution après l’arrêt de la recom-
binaison. Une partie des gènes de la MSY correspondent
à un ensemble de gènes, présents sur la paire autosomale
ancestrale, ayant subi une évolution différentielle sur le
X et le Y [13, 14, 20]. On peut également identifier des
gènes acquis par rétrotransposition ou transposition d’un
autre chromosome, après la divergence initiale du X et
du Y. L’origine génomique de ces gènes est diverse. Deux
gènes sont situés dans une région d’environ 3,4 Mb issue
d’une transposition depuis le X vers le Y, après la diver-
gence de l’homme et du chimpanzé [13, 22]. Deux autres
gènes sont issus d’autosomes pendant l’histoire évolutive
des primates : le gène DAZ est issu du chromosome 3 [23]
et CDY du chromosome 13 [24].
Ultérieurement, les séquences Y, quelle que soit leur
origine, ont évolué globalement de deux fac¸ons différentes
selon leur fonction, mâle-spécifique ou non. Ainsi, une
première portion de la MSY (∼10,2 Mb), la région ampli-
conique, dense en gènes, forme néanmoins un ensemble
fonctionnellement homogène, spécialisé dans la fonction
mâle avec une expression spécifique dans les testicules
[13]. Elle rassemble de longues régions de séquences
dupliquées, le plus souvent agencées selon une structure
en palindrome. Ces gènes correspondent à un ensemble
de familles multigéniques dans lequel le nombre de copies
varie : deux (VCY,XKRY,HSFY,PRY), trois (BPY2) quatre
(CDY,DAZ), six (RBMY) ou même 35 (TSPY) copies [13].
La seconde portion de la MSY regroupe 16 gènes en
copie unique, exprimés dans différents tissus, ainsi que
des pseudogènes, tous homologues des 27 gènes liés au
X, vestiges du temps où le X et le Y constituaient une
paire d’autosomes [13, 21]. La comparaison de séquences
des gènes homologues X et Y permet de reconnaître
des niveaux de divergence très différents (entre 60 et
96 % d’identité) en fonction de leur localisation sur le
X. Entre les paires de gènes X-Y, la divergence augmente
par tranches successives le long du chromosome X en
«strates évolutives »[20]. Actuellement, chez l’homme,
cinq strates évolutives sont reconnues, suggérant au moins
cinq évènements d’arrêt de la recombinaison entre X et Y,
le premier impliquant la région comprenant le gène Sry
[13, 14, 20].
La différenciation X-Y et la spécialisation du Y dans
la fonction mâle sont le résultat d’une longue évolution à
partir d’une paire d’autosomes, suite à la suppression de la
recombinaison. Cela a permis l’acquisition de gènes spé-
cifiques au sexe mâle, mais a également des conséquences
fâcheuses pour le Y.
La dégénérescence du Y
La différenciation progressive du X et du Y est à
l’origine d’un contenu en gènes différent, mais également
d’une structure différente. Le Y est largement hétéro-
chromatique (∼60 %) et présente un grand nombre
de séquences répétées, dont des éléments transposables
comme les LINE ou les séquences Alu [13]. Après environ
150 Ma d’évolution indépendante du X, la région euchro-
matique de la MSY représente 1/6 de la taille du X et n’a
que 1/12 du nombre de gènes [13, 14]. Il apparaît ainsi
qu’en dehors des RPA, le Y humain a perdu 97 % des
gènes initialement présents. De plus, les gènes du Y sont
des homologues sur le X, en version plus ou moins dété-
riorée, quand ils n’ont pas été perdus [13]. En effet, sur
les 27 paires X-Y identifiées, 14 copies Y semblent cor-
respondre à des copies fonctionnelles, codant pour des
isoformes très similaires mais non identiques à celles du
X. Par exemple, les gènes X et Y de l’amélogénine pré-
sentent des différences relatives à leur séquence, au patron
d’épissage et au degré d’expression [25]. Les 13 autres
gènes liés au Y correspondent à des pseudogènes qui
présentent encore une similitude avec leurs homologues
du X relativement importante, en termes de séquence
et de structure (exons-introns) [13]. Le fait que certains
gènes Y soient exprimés pourrait entraîner des différences
de dosage entre les mâles et les femelles à cause de
l’inactivation d’un X chez les femelles. Or, il a été mon-
tré que les gènes Y fonctionnels sont majoritairement
des gènes ayant arrêté de recombiner récemment (strates
4 et 5) et qui échappent, au moins en grande partie, à
l’inactivation du X chez les femelles [26]. Étonnamment,
certains gènes qui échappent à l’inactivation dans ces
strates récentes n’ont pas d’homologue fonctionnel sur le
Y, comme par exemple le gène STS (pseudogène sur le
Y), sans que cela semble poser problème. Les facteurs à
l’origine de l’expression des gènes portés par le X, qu’elle
soit complète, partielle ou inexistante, sont encore très mal
connus. Une meilleure compréhension des mécanismes
de régulation de l’expression des gènes portés par le X
et le Y permettraient probablement de mieux comprendre
l’évolution du déterminisme du sexe et des chromosomes
sexuels.
La faible densité en gènes, la pseudogénéisation et
l’accumulation d’éléments transposables sont les signes
d’une dégénérescence du chromosome Y [27]. Il semble
que les régions du Y qui ont cessé de recombiner en pre-
mier, et notamment la strate 1 comprenant Sry, sont celles
qui montrent les signes de dégénérescence les plus mar-
80 mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 15, n◦1, janvier-février-mars 2013
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

Journal Identification = MTE Article Identification = 0442 Date: March 15, 2013 Time: 4:9 pm
qués, tandis que les seules régions intactes sont les RPA,
qui recombinent encore. La suppression de la recombi-
naison, bien qu’essentielle à l’évolution des chromosomes
sexuels, est donc bien impliquée dans la dégénérescence
du Y. Différents processus peuvent agir sur le Y et entraî-
ner sa dégénérescence [27]. L’absence de recombinaison
entraîne une réduction de la taille efficace des gènes
portés par le Y (un tiers par rapport à ceux du X et un
quart par rapport aux gènes autosomaux) et augmente
de fac¸on considérable l’impact de la dérive génétique
et la fixation d’allèles, y compris d’allèles (faiblement)
délétères. Par ailleurs, les régions du génome non recom-
binantes accumulent des mutations qui, ailleurs dans le
génome, seraient éliminées par sélection naturelle [28].
Ce phénomène, appelé effet Hill-Robertson, explique
la dégénérescence du Y au niveau de leur région non
recombinante : pertes de gènes (pseudogénéisation), accu-
mulation de mutations faiblement délétères (augmentation
du taux de substitutions non synonymes) et de séquences
répétées comme d’éléments transposables [17]. L’absence
de recombinaison aboutit donc, sur le Y, à une diminution
de l’efficacité globale de la sélection naturelle et à une
perte de la diversité génétique en général.
De plus, la MSY humaine est constituée d’une grande
proportion de séquences répétées qui favorisent les réar-
rangements chromosomiques [29], ce qui contribue à
l’érosion du Y. Chez l’homme, les remaniements chro-
mosomiques du Y sont fréquents et à l’origine d’un
important polymorphisme de structure [30]. Ces remanie-
ments peuvent entraîner la délétion de grandes portions de
la MSY comprenant des gènes impliqués dans la fertilité
et donc potentiellement fortement sélectionnés. Ces délé-
tions de la MSY peuvent être pathologiques (infertilité [31])
mais peuvent aussi exister sans impact clairement défini
sur la fertilité. Ainsi, la délétion gr/gr couvrant 1,6 Mb
présente une prévalence mesurée dans les études de cas
d’environ 5 %, apparaît comme un facteur de risque pour
l’infertilité masculine, notamment oligozoospermie en
Europe, même si la relation entre présence de la délétion
et infertilité dépend de l’ethnicité et de la région géogra-
phique [32]. Étonnamment, des Y amputés d’une région
de 1,8 Mb incluant 12 gènes à fonction mâle-spécifique
(délétion b2/b3, proche en termes de structure de la délé-
tion gr/gr) dont la moitié des gènes de la région AZFc,
cruciale pour une spermatogenèse normale, sont pourtant
largement distribués dans le nord de l’Eurasie [33].
Un facteur supplémentaire contribue à la dégénéres-
cence duY:lenombre de divisions cellulaires plus élevé
dans la lignée germinale mâle que femelle, à l’origine d’un
taux de mutation plus élevé sur le Y. L’impact de ce facteur
sur la dégénérescence du Y a été validé grâce à la compa-
raison des systèmes XY et ZW. En effet, alors que les Y
et W dégénèrent dans ces deux systèmes, le Y est présent
uniquement chez les mâles et le W chez les femelles. Il a
été montré que le taux de mutation n’est pas plus élevé sur
le W que sur les autosomes [34]. Chez les grands singes
et l’homme, l’importance de cette évolution moléculaire
par les mâles (male-driven evolution), c’est-à-dire le ratio
du taux de mutation mâle-femme, a été estimée de l’ordre
de 5 [35].
Ainsi, la suppression de la recombinaison a permis la
spécialisation du Y mais elle a également entraîné la perte
de gènes, l’accumulation de mutations et de séquences
répétées conduisant à la dégénérescence de ce chro-
mosome. Les mécanismes moléculaires à l’origine de la
dégénérescence du Y sont toujours actuellement en œuvre
(taux de mutation plus élevé, sélection moins efficace,
dérive génétique accrue) et il n’y a pas de raison pour
que cela change. L’étude des chromosomes Y et W dans
différentes lignées animales (vertébrés ou non) et même
chez les plantes montre une dégradation inexorable. La
dégénérescence du chromosome Y humain ne fait aucun
doute, mais alors cette dégradation va-t-elle continuer
jusqu’à ce que le Y disparaisse ou va-t-on atteindre un
point d’équilibre ? Beaucoup de débats ont lieu autour de
cette question sensible [par exemple voir 36] et il existe
des arguments pour et contre.
Le chromosome Y humain
va-t-il disparaître ?
Chez l’homme, la dégradation des gènes du Y a été
très rapide : perte de plus de 1 000 gènes en 150 Ma, soit
en moyenne 6,6 gènes par Ma. Si on extrapole, avec une
perte linéaire, le Y humain devrait disparaître dans envi-
ron 5 Ma [16, 36]. Cependant, il semble peu probable que
le taux de dégradation soit linéaire puisque la suppression
de la recombinaison a été progressive, modifiant ainsi le
nombre de gènes potentiellement affectés par la dégrada-
tion (balance entre gènes recrutés dans la MSY et gènes
déjà perdus), modifiant le modèle de dégradation qui cor-
respondrait alors à une sinusoïdale [16]. De plus, le Y
humain est composé de deux régions distinctes (ancestrale
et ajoutée après la divergence des marsupiaux et placen-
taires), le modèle de dégradation du Y humain correspond
plus vraisemblablement à une double sinusoïdale rendant
difficile une prédiction sur la date de l’extinction du Y [16],
mais augmentant probablement celle-ci.
Récemment, la publication des séquences complètes
du Y du chimpanzé [37] et du macaque rhésus [38] a
permis la comparaison avec le Y humain. Ces données
réfuteraient la disparition du Y et suggèrent que le Y pour-
rait être maintenu indéfiniment grâce à la pression de
sélection exercée sur les gènes mâle-spécifiques [36, 38].
En effet, il apparaît que depuis la séparation de l’homme et
du chimpanzé, soit environ 6 Ma, la MSY humaine n’aurait
perdu aucun gène : aucun gène présent chez le chimpanzé
n’a été perdu chez l’homme [37, 39]. La comparaison des
Y humain et du rhésus, divergeant depuis 25 Ma, montre
mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 15, n◦1, janvier-février-mars 2013 81
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

Journal Identification = MTE Article Identification = 0442 Date: March 15, 2013 Time: 4:9 pm
Mini-revue
que le contenu génique a été largement maintenu sur cette
période de l’évolution du Y et aucune perte de gène n’est
visible au niveau des strates évolutives les plus anciennes
[38]. Les pertes de gènes ne se sont produites que dans
la strate la plus récente, formée juste avant la séparation
de l’homme et du rhésus. Sur les sept gènes présents sur
cette strate au départ, un seul est conservé chez l’homme
et deux seulement chez le rhésus. Ces différentes don-
nées suggèrent que la perte a été initialement rapide avant
de ralentir de manière importante, voire potentiellement
de s’arrêter dans les strates les plus anciennes [38]. De
manière intéressante, il semblerait que le chimpanzé ait
souffert d’une perte de gènes importante : six gènes depuis
sa séparation de la lignée humaine, suggérant une accé-
lération récente de la dégradation chez les chimpanzés
[37, 39].
La comparaison du Y de ces trois espèces de pri-
mates montre également une dynamique particulière des
séquences ampliconiques, très différente de celle identi-
fiée pour les paires de gènes X-Y. Les séquences amplico-
niques ont subi une restructuration drastique dans les trois
lignées, suivie d’une évolution indépendante et complète-
ment différente [37, 38]. D’abord, la proportion de la MSY
euchromatique occupée par les séquences ampliconiques
est très différente : 45 % chez l’homme, 57 % chez le chim-
panzé et5%chezlemacaque. Le chimpanzé présente
donc une région ampliconique bien plus grande, mais
aussi bien plus complexe, avec souvent plus de copies
de gènes que chez l’homme [37, 38]. Étant donné la
divergence de ces espèces, l’évolution des régions ampli-
coniques a été rapide et pourrait résulter d’une sélection
positive favorisant les gènes impliqués dans la sperma-
togenèse [38]. La nature même de ces régions riches
en séquences répétées, favorisant les remaniements, peut
également contribuer à leur évolution rapide.
Les séquences ampliconiques agencées en palindrome
sont très similaires en termes de séquences (>99,9 %
identité) [13]. Le mécanisme de conversion génique intra-
Y maintient un haut degré d’identité entre les copies
de gènes et peut ainsi protéger l’intégrité de gènes cri-
tiques pour la spermatogenèse [40]. De récentes études
basées sur des modèles de génétique des populations et
des simulations montrent que même de faibles niveaux
de conversion génique sont suffisants pour maintenir
l’intégrité des copies Y et empêcher la dégénérescence
du Y dans ces régions [41]. Cependant, le processus de
conversion génique n’est pas directionnel et une copie
fonctionnelle peut être convertie en une copie mutée.
L’étude des palindromes humains a d’ailleurs révélé
29 copies du gène RBMY dont seulement six sont actives
[13]. La sélection naturelle reste donc indispensable pour
éliminer les Y de la population contenant des copies
mutées de gènes impliqués dans la fertilité.
En revanche, des gènes en copie unique avec une
fonction essentielle, comme Sry, ne peuvent pas être
restaurés par le mécanisme de conversion génique. Ils
doivent cependant être soumis à de très fortes pressions
de sélection. L’étude du polymorphisme chez l’homme
(∼100 individus) a montré une remarquable conservation
de séquences pour les gènes Y ayant un homologue sur le
X suggérant une importante sélection purificatrice [42].
Les gènes sous forte pression de sélection doivent être
extrêmement difficiles à perdre, ce qui fait que certains
modèles proposent que le Y humain puisse être main-
tenu grâce à une sélection positive sur le long terme [36].
Pourtant, il semble que des gènes fortement sélectionnés
puissent être perdus, si leur fonction est remplacée par
d’autres. Par exemple chez les marsupiaux, les gènes ATRX
et ATRY montrent des expressions spécifiques, respecti-
vement dans les tissus somatiques et dans les gonades.
Malgré cela, le gène ATRY a été perdu chez les placen-
taires où ATRX est exprimé également dans les gonades,
suggérant que la fonction du gène ATRY a été reprise par
son homologue X [43]. Le recrutement de copies autoso-
males pourrait faciliter la perte des gènes Y. Par exemple, il
existe dans le génome humain, outre les multiples copies
du facteur de spermatogenèse RBMY et la copie homo-
logue RBMX, plusieurs copies autosomales, dont l’une est
active au niveau des testicules [44] et pourrait être recrutée
en replacement de RBMY.
Étant donné la nature stochastique des facteurs en
jeu (remaniements chromosomiques et délétions ; acqui-
sition et/ou duplication de gènes avantageux pour la
fonction mâle), l’avenir du Y est peu prévisible et soumis
au facteur chance ; il est donc peu probable qu’un état
d’équilibre stable soit actuellement atteint [16]. Il suffirait
de quelques substitutions de fonctions pour uniquement
quelques gènes pour rendre le Y humain totalement redon-
dant et permettre sa perte complète [16]. Ainsi, selon J.
Graves, la question serait de savoir non pas si le chro-
mosome Y disparaîtra mais quand il disparaîtra [36]. Un
argument de poids est la disparition du Y chez de nom-
breuses espèces animales. Les espèces de drosophiles
procurent des exemples spectaculaires de disparition du
Y, avec maintien des mâles [45, 46]. Chez les mam-
mifères, la perte du Y voudrait-elle dire la disparition
du sexe mâle et une reproduction du type parthénoge-
nèse ? Rien n’est moins sûr... Chez les mammifères,
le phénomène d’empreinte parentale rend obligatoire la
présence d’un génome mâle et d’un génome femelle.
Chez les mammifères, le système de déterminisme XX/XY
est particulièrement conservé, pourtant il existe quelques
exceptions. En effet, d’autres systèmes de déterminisme
du sexe existent, tous chez des rongeurs, et dérivent du
système ancestral XX/XY. Ils se regroupent en différentes
catégories, impliquant ou non la disparition du Y.
On a pu identifier des espèces où les femelles sont X0
(1 seul X) et les mâles sont XY, comme chez le campa-
gnol Microtus oregoni [47]. Ce système est très proche du
XX/XY ancestral, où seul un X disparaît chez les femelles.
82 mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 15, n◦1, janvier-février-mars 2013
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.
 6
7
8
9
6
7
8
9
1
/
9
100%