Exploration de la mémoire épisodique par l`imagerie fonctionnelle

Article original
Article original
Exploration
de la mémoire épisodique
par l’imagerie fonctionnelle :
données chez l’adulte
Sophie Dupont
Unité d’Épileptologie, 41 bd de l’Hôpital, 75013 Paris, France
Résumé. Historiquement, la mémoire a été reliée au circuit de Papez. L’imagerie fonctionnelle que ce soit la
tomographie par émission de positons ou ultérieurement l’imagerie par résonance magnétique fonctionnelle a
permis de faire évoluer ces concepts historiques. Les premières études réalisées chez des sujets volontaires sains, ont
démontré in vivo la possibilité d’activer les formations hippocampiques et parahippocampiques dans des paradigmes
de mémoire visuelle ou verbale, confirmant ainsi les connaissances anatomiques issues des corrélations anatomo-
cliniques mais ont également mis en exergue le rôle joué par d’autres régions cérébrales, notamment préfrontales et
pariétales. L’anatomie de la mémoire doit chez le sujet sain s’envisager en termes de circuits corticaux au sein
desquels certaines régions, comme les régions temporales médiales, pourraient jouer un rôle déterminant, mais non
exclusif. Chez le sujet épileptique, la plasticité de ces circuits qui potentiellement sous-tend les possibilités de
récupération fonctionnelle reste à explorer.
Mots clés :IRMf, effet BOLD, réorganisation, cartographie cognitive
Abstract. Exploration of episodic memory by fonctional imaging: results by adults
Historically, the memory was presumed to be subserved by the Papez circuit. Functional imaging, either positron
emission tomography or functional MRI, has helped to change this historical concepts. Recent findings have
converged on the observation that hippocampal or parahippocampal regions could be consistently activated in
verbal or visuospatial memory paradigms. Furthermore, these functional neuroimaging studies have also stressed
the importance of the prefrontal and parietal cortical activations in such memory processes. These results show that
memory is subserved by a more widely distributed cortical network than expected. The plasticity of these circuits
must be now explored in epileptic patients.
Key words:fMRI, bold effect, reorganization, cognitive cartography
L’IRMf représente un outil d’avenir pour
l’exploration préchirurgicale des patients épi-
leptiques. Différents champs d’investigation
sont explorés de façon parallèle : la localisation
du foyer épileptogène grâce à l’enregistrement
combiné de l’EEG sous l’IRMf, la délimitation
fonctionnelle des aires sensitives et motrices
permettant la planification des résections cor-
ticales adjacentes et surtout la localisation de
fonctions cognitives telles que le langage ou la
mémoire, pouvant représenter une alternative
au test de Wada voire même un outil prédictif
indépendant du devenir neuropsychologique
postopératoire.
Dans cet article, nous envisagerons succes-
sivement l’apport de l’IRMf dans l’exploration
de la mémoire épisodique chez le sujet sain et
chez le sujet épileptique.
53 E
´pilepsies, vol. 18, Numéro spécial, septembre 2006
E
´pilepsies 2006 ; 18 (Numéro spécial) : 53-6
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

Mémoire, neuropsychologie
et imagerie fonctionnelle
Il existe, on le sait, différents types de mémoire sous-tendus
par des réseaux neuronaux différents. Il est impossible d’être ici
exhaustif et d’envisager successivement l’apport de l’imagerie
fonctionnelle dans tous ces différents types de mémoire. Nous
nous contenterons donc de l’exemple de la mémoire épisodi-
que, dont le dysfonctionnement est le plus souvent souligné
dans l’épilepsie, notamment temporale interne.
Mémoire chez le sujet sain
Hippocampe et mémoire épisodique : vérification
et consolidation des données neuropsychologiques
Les études en imagerie fonctionnelle ont permis d’attester la
réalité et l’importance de l’activation hippocampique dans les
systèmes de mémoire épisodique, importance qui avait déjà été
suggérée par les données cliniques (patient HM, tests neuropsy-
chologiques) et l’expérimentation animale.
C’est initialement dans des paradigmes de mémoire visuo-
spatiale qu’une activation hippocampique a été obtenue de
façon reproductible et fiable (Stern et al., 1996). Ultérieurement,
des activations hippocampiques ont également été décrites
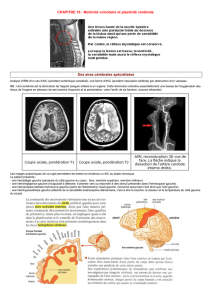
dans des tâches de mémoire verbale. Golby et al. ont clairement
démontré la notion de spécialisation hippocampique dans le
traitement des informations verbales et visuo-spatiales (Golby et
al., 2001) (figure 1). L’hippocampe droit (ou non dominant) est
ainsi préférentiellement impliqué dans les tâches de mémoire
visuo-spatiale alors que l’hippocampe gauche (ou dominant)
intervient préférentiellement dans les processus intéressant la
mémoire verbale. Gabrieli et al. ont pour leur part suggéré qu’il
existait en outre une implication différentielle des parties anté-
rieures et postérieures de l’hippocampe selon la tâche mnésique
réalisée (Gabrieli et al., 1997). Ils avaient ainsi détecté une
activation de l’hippocampe postérieur et du gyrus parahippo-
campique dans une tâche d’encodage de scènes visuelles com-
plexes et une activation de la partie antérieure de l’hippocampe
et du subiculum au cours d’une tâche de reconnaissance des
figures précédemment encodées. L’hypothèse avancée pour ex-
pliquer cette dichotomie postéro-antérieure était que l’enco-
dage de mémoire épisodique nécessitait une distinction du
caractère familier ou non du stimulus et que cette distinction
faisait appel principalement au cortex parahippocampique,
connu pour recevoir de fortes projections des zones visuelles
d’intégration. A l’inverse, la tâche de rappel qui faisait appel aux
informations stockées dans d’autres zones néocorticales, néces-
sitait l’activation de l’hippocampe antérieur et surtout du subi-
culum, principale voie de sortie des informations hippocampi-
ques vers les régions corticales et sous-corticales. Cette
dichotomie antéro-postérieure a été retrouvée dans d’autres
études IRMf de la littérature (Bernard et al., 2004) mais reste
controversée aux vues des résultats d’une méta-analyse TEP
donnant des résultats inverses (Lepage et al., 1998).
Par ailleurs, les études en IRMf réalisées chez le sujet sain
s’intéressent spécifiquement au rôle exact joué par l’hippo-
campe dans les étapes et processus de la mémorisation et à ce
titre, l’hippocampe semble jouer un rôle prépondérant à bien
des niveaux :
–
rôle dans la détection de la nouveauté : l’hippocampe s’active-
rait spécifiquement dans la détection du caractère non familier
des stimuli, étape préliminaire de l’encodage proprement dit ;
–
rôle dans le rappel conscient des informations (Wheeler et al.,
2004) ;
–
consolidation de la trace mnésique.
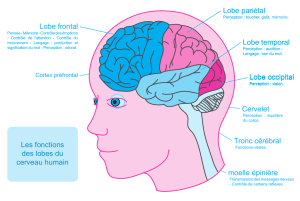
Imagerie fonctionnelle et mémoire épisodique :
ouverture vers le cerveau
L’un des principaux apports de l’imagerie fonctionnelle est
également d’avoir démontré que la mémoire épisodique ne se
réduisait pas qu’au seul système hippocampo-diencéphalique et
donc au circuit décrit par Papez dans les années trente, et notam-
ment d’avoir mis l’accent sur le rôle joué par le cortex parahip-
pocampique (au sens général du terme), le lobe frontal et le lobe
pariétal dans le fonctionnement de la mémoire épisodique.
L’activation du cortex parahippocampique est retrouvée de
manière consistante dans la plupart des paradigmes d’IRMf
s’intéressant à la mémoire épisodique (Stern et al., 1996). Diffé-
rentes régions parahippocampiques semblent préférentielle-
ment s’activer : le cortex entorhinal, le cortex périrhinal, voire
même le subiculum (Zeineh et al., 2000). Les limites anatomi-
ques de ces activations ont été définies grâce à de nouvelles
méthodes d’IRMf utilisant à la fois une IRM à haut champ et une
technique sophistiquée de segmentation et déplissage du cortex
(Zeineh et al., 2005). Un article récent d’Eldridge et al. (Eldridge
et al., 2005) suggère même que seul le subiculum serait activé
pendant les phases de rappel alors que le gyrus denté et les
champs CA2, CA3 seraient spécifiquement activés pendant
l’encodage. Ces constatations rejoignent la théorie de la dicho-
tomie antéro-postérieure hippocampique selon un axe rappel-
Figure 1
.
Activation hippocampique (données personnelles).
S. Dupont
54
E
´pilepsies, vol. 18, Numéro spécial, septembre 2006
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

encodage de Gabrieli, 1997). Le rôle joué par le cortex parahip-
pocampique dans les processus de mémorisation demeure plus
hypothétique :
–
passage obligé d’entrée et de sortie pour les informations
allant à l’hippocampe ?
–
rôle propre de stockage des informations ?
Pour ce qui est du lobe frontal, les études en tomographie
par émission de positons ont été pionnières dans ce domaine et
doivent être rapportées ici pour mieux comprendre les résultats
obtenus ultérieurement en IRMf. En 1994, Kapur et al. ont
étudié en TEP la mémoire épisodique chez 12 sujets volontaires
sains (Kapur et al., 1995). Ils ont mis en évidence une activation
intense du cortex inférieur préfrontal gauche (aires de Brod-
mannn 45, 46, 47 et 10) dans la tâche d’encodage et une
activation du cortex frontal dorsolatéral droit (aires de Brod-
mannn 10, 46 et 9) dans la tâche de rappel. Cette asymétrie dans
l’activation du cortex préfrontal dans des tâches d’encodage et
de rappel de mémoire épisodique a été conceptualisée sous le
terme de modèle HERA (Hemispheric encoding/retrieval asymmetry
model) (Nyberg et al., 1996). Ce modèle HERA a le mérite d’avoir
attiré l’attention sur le rôle joué par le cortex frontal dans les
processus de mémorisation même si la dichotomie gauche-
droite selon un axe encodage-rappel n’a pas été toujours vérifiée
dans la littérature. Néanmoins, la plupart des études en IRMf
retrouvent de consistantes activations du cortex préfrontal droit
dans les tâches de rappel épisodique.
Les explications avancées pour expliquer la mise en jeu du
cortex frontal dans les processus de mémorisation épisodique
sont multiples et fonction de l’aire frontale concernée (Fletcher
et al., 2001) :
–
rôle de maintien et mise à jour des informations pour le cortex
ventrolatéral ;
–
rôle de sélection, manipulation et contrôle des informations
pour le cortex dorsolatéral ;
–
rôle de sélection des processus, des objectifs à atteindre et des
moyens d’y parvenir pour le pôle frontal latéral.
Par ailleurs, des activations reproductibles du cortex pariétal
gauche et droit et du précunéus ont également été retrouvées
dans des tâches d’encodage et de rappel (Konishi et al., 2000).
Ces activations sont mises en parallèle avec le rôle connu du
cortex pariétal dans la manipulation spatiale des informations
et leur sélection qui pourraient constituer une étape importante
des processus de mémorisation.
Mémoire chez les patients épileptiques
Remplacement du test de Wada
Il existe peu d’études d’IRMf ayant exploré la mémoire chez
le patient épileptique.
Les cartographies cognitives explorent essentiellement le
langage et la mémoire et reposent à l’heure actuelle sur le test de
Wada considéré comme le « gold standard » en la matière.
Des études menées en comparaison directe avec le test de
Wada ont montré une concordance des résultats quant à la
latéralisation du langage avec en outre en IRMf de nombreuses
informations complémentaires sur la localisation précise des
différentes zones impliquées dans le langage (Binder et al. 1995 ;
Binder et al., 1996 ; Thivard et al., 2005).
Pour ce qui est de la mémoire, seules quatre études (Deblaere
et al., 2005 ; Detre et al., 1998 ; Golby et al., 2002 ; Rabin et al.,
2004) se sont intéressées directement au remplacement du test de
Wada mémoire par l’IRMf en comparant les résultats des deux
procédures chez les mêmes patients souffrant d’épilepsie tempo-
rale pharmacorésistante candidats à la chirurgie. Les deux pre-
mières études (Detre et al., 1998 ; Golby et al., 2002) réalisées sur
de faibles cohortes de patients (respectivement 10 et 9 patients)
retrouvaient une bonne concordance entre les deux méthodes
avec pour l’IRMf comparée au Wada une sensibilité oscillant entre
90 et 100 % et une spécificité entre 88 % et 100 %. La troisième
étude (Rabin et al., 2004) qui a été réalisée sur un plus grand
nombre de patients épileptiques (35) montrait une corrélation
positive entre les activations temporales médiales et les ratios
d’asymétrie obtenus au Wada sur une analyse de groupe, mais les
corrélations individuelles n’étaient pas disponibles. La quatrième
étude (Deblaere et al., 2005) réalisée elle aussi sur une cohorte plus
vaste de 18 patients a retrouvé, quant à elle, des résultats plus
discordants avec uniquement une bonne concordance entre le
Wada et les activations obtenues en IRMf uniquement pour les
patients temporaux droits (n = 10) mais pas pour les gauches.
Hippocampe et mémoire épisodique : nouvelles hypothèses
On le voit donc, au vu des résultats de la littérature, le
remplacement du Wada mémoire par l’IRMf n’est pas encore
totalement validé. Ce, d’autant plus que se dégage actuellement
un courant distinct d’évaluation de la mémoire cherchant à
s’affranchir de la comparaison au test de Wada, de plus en plus
remis en question (Abou-Khalil et al., 2002).
De nombreuses études d’imagerie fonctionnelle s’intéressent
actuellement à la valeur prédictive isolée des activations hippo-
campiques en préopératoire, sans référence au test de Wada.
Pour prédire un éventuel déficit mnésique postopératoire,
deux données vont alors être prises en compte : la valeur fonc-
tionnelle résiduelle de l’hippocampe épileptique (capacité fonc-
tionnelle), et la valeur fonctionnelle de l’hippocampe controla-
téral ainsi que sa capacité à reprendre en charge la fonction
défaillante de l’autre hippocampe (réserve fonctionnelle) (Che-
lune et al., 1995). En clair et de façon logique, si l’hippocampe
épileptique que l’on désire enlever chirurgicalement s’active
bien en imagerie fonctionnelle, c’est de mauvais pronostic
puisque l’on va ôter une structure encore fonctionnellement
capable ; si l’hippocampe épileptique que l’on désire enlever
chirurgicalement s’active peu ou pas et qu’à l’inverse, l’hippo-
campe controlatéral s’active, c’est de bon pronostic puisque l’on
ôte une structure non fonctionnelle et que la structure paire
peut prendre le relais (phénomène de plasticité ?). Ces hypothè-
ses ont déjà été vérifiées en tomographie de positons (Henke et
al., 2003) et en IRMf (Richardson et al., 2004).
Réorganisation des réseaux cortico-hippocampaux :
vers la plasticité ?
Des études ont été menées sur la réorganisation des réseaux
cortico-hippocampiques sous-tendant la mémoire chez les pa-
55 E
´pilepsies, vol. 18, Numéro spécial, septembre 2006
Exploration de la mémoire épisodique par l’imagerie fonctionnelle : données chez l’adulte
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

tients épileptiques souffrant d’une sclérose hippocampique
droite ou gauche (Dupont et al., 2000 ; Dupont et al., 2001 ;
Dupont et al., 2002). Les résultats ont montré que les patients
souffrant d’une épilepsie de la face médiale gauche activaient
des réseaux cérébraux qui différent de ceux des témoins princi-
palement en regard du cortex temporo-médial (gyrus parahip-
pocampique et hippocampe) et du cortex préfrontal dorsolaté-
ral ; ces différences dans les patterns d’activations étant censés
refléter soit un dysfonctionnement soit une réorganisation des
réseaux neuronaux normaux sous-tendant la mémoire. La plas-
ticité hippocampique est à l’heure actuelle encore assez mal
évaluée en imagerie fonctionnelle. Les données issues des obser-
vations post-opératoires (cf. chapitre correspondant) devraient
à l’avenir nous apporter des renseignements pertinents.
Conclusion
L’IRMf permet actuellement de visualiser les régions céré-
brales activées par des tâches mnésiques. La question posée
actuellement par beaucoup d’auteurs est celle du lien entre la
neuropsychologie et l’imagerie fonctionnelle (Shallice et al.,
2003). Les complexes modèles neuropsychologiques peuvent-
ils être mis en parallèle avec la vision brute et instantanée de
l’activation cérébrale apportée par l’imagerie fonctionnelle ?
Certes non, beaucoup de questions restent posées par les images
d’activation cérébrale. Comment être certain que les circuits
visualisés correspondent bien à la tâche réalisée ? Comment
contrôler l’efficience de la tâche et les tâches intercurrentes?
Quel lien entre le degré d’activation et le degré d’efficience de la
tâche effectuée ? Quel lien dynamique et temporel entre les
régions cérébrales activées ? Quid des zones désactivées pen-
dant ces tâches ? Des protocoles d’imagerie et des techniques
d’analyse de plus en plus complexes élaborés en collaboration
avec les neuropsychologues et les méthodologistes tentent de
répondre à ces questions. Gageons que dans l’avenir, les deux
approches neuropsychologiques et fonctionnelles sauront se
compléter et s’enrichir mutuellement.
M
Références
Abou-Khalil BW, Schlaggar BL. Is it time to replace the Wada test?
Neurology 2002 ; 59 : 160-1.
Bernard F, Bullmore E, Graham K, Thompson S, Hodges J, Fletcher P.
The hippocampal region is involved in successful recognition of both
remote and recent famous faces. Neuroimage 2004 ; 22 : 1704-14.
Binder JR, Rao SM, Hammeke TA, et al. Lateralized human brain
language systems demonstrated by task subtraction functional magnetic
resonance imaging. Arch Neurol 1995 ; 52 : 593-601.
Binder JR, Swanson SJ, Hammeke TA, et al. Determination of language
dominance using functional MRI : a comparison with the Wada test.
Neurology 1996 ; 46 : 978-84.
Chelune G. Hippocampal adequacy versus functional reserve :
predicting memory functions following temporal lobectomy. Arch Clin
Neuropsychol 1995 ; 10 : 413-32.
Deblaere K, Backes W, Tieleman A, et al. Lateralized anterior
mesiotemporal lobe activation : semirandom functional MR imaging
encoding paradigm in patients with temporal lobe epilepsy--initial
experience. Radiology 2005 ; 236 : 996-1003.
Detre JA, Maccotta L, King D, et al. Functional MRI lateralization of
memory in temporal lobe epilepsy. Neurology 1998 ; 50 : 926-32.
Dupont S, Van de Moortele P, Samson S, et al. Episodic memory in left
temporal lobe epilepsy : a functional MRI study. Brain 2000 ; 123 : 1722-32.
Dupont S, Samson Y, Van de Moortele P, et al. Verbal memory
consolidation : a functional MRI study in healthy volunteers and patients
with left medial temporal lobe epilepsy. Neuroimage 2001 ; 14 : 995-1003.
Dupont S, Samson Y, Van de Moortele P, et al. Bilateral hippocampal
dysfunction evidence in unilateral medial temporal lobe epilepsy. J Neurol
Neurosurg Psychiatry 2002 ; 73 : 478-85.
Eldridge L, Engel S, Zeineh M, Bookheimer S, Knowlton B. A
dissociation of encoding and retrieval processes in the human
hippocampus. J Neurosci 2005 ; 25 : 3280-6.
Fletcher P, Henson R. Frontal lobes and human memory : insights
from functional neuroimaging. Brain 2001 ; 124 : 849-81.
Gabrieli JDE, Brewer JB, Desmond JE, Glover GH. Separate neural
bases of two fundamental memory processes in the human medial
temporal lobe. Science 1997 ; 276 : 264-6.
Golby A, Poldrack R, Brewer J, et al. Material-specific lateralization in
the medial temporal lobe and prefrontal cortex during memory encoding.
Brain 2001 ; 124 : 1841-54.
Golby A, Poldrack R, Illes J, Chen D, Desmond J, Gabrieli J. Memory
lateralization in medial temporal lobe epilepsy assessed by functional MRI.
Epilepsia 2002 ; 43 : 855-63.
Henke K, Treyer V, Weber B, et al. Functional neuroimaging predicts
individual memory outcome after amygdalohippocampectomy.
Neuroreport 2003 ; 14 : 1197-202.
Kapur S, Craik FIM, Jones C, Brown GM, Houle S, Tulving E.
Functional role of the prefrontal cortex in retrieval of memories : a PET
study. Neuroreport 1995 ; 6 : 1880-4.
Konishi S, Wheeler M, Donaldson D, Buckner R. Neural correlates of
episodic retrieval success. Neuroimage 2000 ; 12 : 276-86.
Lepage M, Habib R, Tulving E. Hippocampal PET activations of
memory encoding and retrieval : the HIPER model. Hippocampus 1998 ; 8 :
313-22.
Nyberg L, Cabeza R, Tulving E. PET studies of encoding and retrieval :
the HERA model. Psychon Bull Rev 1996 : 3.
Rabin M, Narayan V, Kimberg D, et al. Functional MRI predicts post-
surgical memory following temporal lobectomy. Brain 2004 ; 127 : 2286-98.
Richardson M, Strange B, Thompson P, Baxendale S, Duncan J,
Dolan R. Pre-operative verbal memory fMRI predicts post-operative
memory decline after left temporal lobe resection. Brain 2004 ; 127 :
2419-26.
Shallice T. Functional imaging and neuropsychology findings : how
can they be linked? Neuroimage 2003 ; 20 : S146-S154.
Stern CE, Corkins S, Gonzales RG, et al. The hippocampal formation
participates in novel picture encoding : evidence from functional magnetic
resonance imaging. Proc Natl Acad Sci USA 1996 ; 93 : 8660-5.
Thivard L, Hombrouck J, du Montcel S, et al. Productive and
perceptive language reorganization in temporal lobe epilepsy. Neuroimage
2005 ; 24 : 841-51.
Wheeler M, Buckner R. Functional-anatomic correlates of
remembering and knowing. Neuroimage 2004 ; 21 : 1337-49.
Zeineh M, Engel S, Bookheimer S. Application of cortical unfolding
techniques to functional MRI of the human hippocampal region.
Neuroimage 2000 ; 11 : 668-83.
Zeineh M, Engel S, Thompson P, Bookheimer S. Dynamics of the
hippocampus during encoding and retrieval of face-name pairs. Science
2005 ; 299 : 3280-6.
S. Dupont
56
E
´pilepsies, vol. 18, Numéro spécial, septembre 2006
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.
1
/
4
100%