La cellule de Langerhans - iPubli

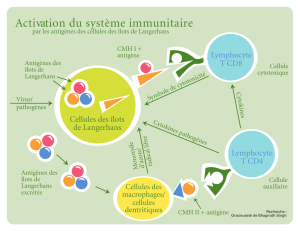
La cellule de Langerhans
D




 6
7
8
9
6
7
8
9
1
/
9
100%