PHYSIOLOGIE DU MUSCLE. {par Jean

PHYSIOLOGIE DU MUSCLE.
{par Jean-Pierre GOUSSARD}
Introduction. 2
1) Anatomie du muscle squelettique. 2
1.1) Composition chimique du muscle. 3
2) Ultrastructure du muscle squelettique. 3
2.1) Le sarcomère. 3
2.1.1) Caractéristique des myofilaments. 4
2.2) Le système tubulaire intracellulaire. 6
2.2.1) Fonction des tubules transverses. 7
3) Théorie de l'interdigitation. 7
3.1) Mécanisme moléculaire de l'interdigitation. 9
3.1.1) Action mécaniques des ponts. 9
3.2) Mécanisme de l'interdigitation. 10
4) Myotopologie. 12
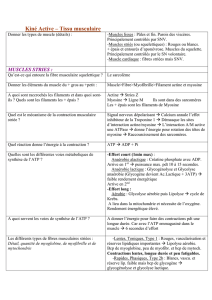
4.1) Caractéristiques des fibres ST 12
4.2) Caractéristiques des fibres FT 13
4.3) Modifications des fibres Musculaires. 14
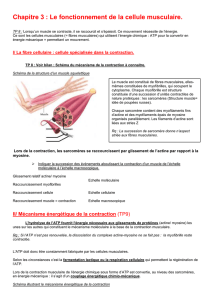
5) Mécanisme de la contraction musculaire. 15
5.1) Secousses musculaires. 15
5.2) Sommation et tétanos. 16

J-P. Physiologie du muscle.
-2-
Introduction.
Les muscles représentent 40 à 50% du poids total du corps humain. La
fonction principale du processus contractile consiste à produire des forces
et des mouvements en se contractant. La contraction des muscles
squelettiques, placée sous le contrôle du système nerveux somatique,
permet non seulement à l'homme de se déplacer dans son environnement,
de le contrôler, de le modifier, de saisir et de déplacer des objets mais aussi
de participer à un rôle important dans la régulation thermique corporelle.
Chez l'homme, il existe trois types de cellules musculaires :
- le muscle lisse, généralement placé sous le contrôle du système nerveux
autonome assure le fonctionnement de la vie végétative.
- le muscle cardiaque, lui aussi placé sous le contrôle du système nerveux
autonome assure la contraction du myocarde.
- Le muscle squelettique, qui permet avant tout les interactions entre le
corps et son environnement.
Bien qu'ils existent des différences importantes de structures, de propriétés
mécaniques et de mécanisme de contrôle entre ces trois types de muscles,
la production de force est la même pour chacun d'eux.
1) Anatomie du muscle squelettique.
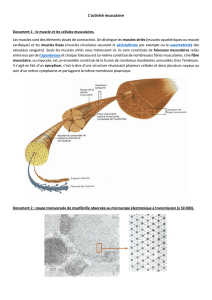
Un muscle squelettique est constitué de milliers de cellules musculaires
cylindriques nommées fibres musculaires. Une seule cellule musculaire
constitue une fibre musculaire. Chaque fibre musculaire squelettique est
un cylindre ayant un diamètre de 10 à 100 microns et une longueur qui
peut atteindre 30 cm. Ces longues et fines fibres multinucléées sont
parallèles les unes aux autres et la force de contraction s'applique dans le
sens du grand axe de la fibre.
Chaque fibre est enveloppée et séparée des autres par une mince couche
de tissu conjonctif, l'endomysium. Une autre couche de tissu conjonctif,
le périmysium, enveloppe jusqu'à 150 fibres pour former un faisceau. Un
fascia de tissu conjonctif fibreux, l'épimysium, enveloppe l'ensemble du
muscle. Cette gaine protectrice se referme aux extrémités et s'unit aux
autres enveloppes du tissu intramusculaire pour former le tissu
conjonctif dense et fort des tendons. Les tendons réunissent chaque
extrémité du muscle à la membrane externe des os, le périoste.
Le sarcolemme situé sous l'endomysium entoure chaque fibre musculaire.
Cette fine membrane élastique délimite le contenu cellulaire de la fibre.
Le protoplasme acqueux ou sarcoplasme de la cellule renferme des
protéïnes contractiles, des enzymes, des particules de graisse et de
glycogène, des noyaux et plusieurs autres organites spécialisés.

J-P. Physiologie du muscle.
-3-
1.1) Composition chimique du muscle.
Le muscle squelettique est constitué d'environ 75% d'eau et 20% de
protéïne ; le reste consiste en sels inorganiques et autres substances
comme des phosphates riches en énergie, de l'urée, de l'acide
lactique et diverses minéraux, des enzymes et pigments, des ions, des
acides aminés, des graisses et des sucres.
Les protéïnes musculaires les plus abondantes sont la myosine,
l'actine et la tropomyosine. Elles représentent respectivement 52%,
23% et 15% du contenu protéïque musculaire. En outre, il faut
environ 700 mg de myoglobine par 100 g de tissu musculaire.
2) Ultrastructure du muscle squelettique.
Chaque fibre musculaire est constituée de petites unités fonctionnelles
parallèles au grand axe de la fibre : les fibrilles. Ces fibrilles, encore
appelées myofibrilles, ont un diamètre d'environ un micron ; elles sont
aussi constituées de sous unités encore plus petites : les filaments ou
myofilaments, également parallèle au grand axe de la myofibrille.
Les myofilaments sont principalement composés de deux protéïnes,
l'actine et la myosine. Six autres protéïnes jouent un rôle structural ou
sont impliquées dans l'interaction des protéïnes filamenteuses au cours
de la contraction musculaire. Ce sont :
- la tropomyosine, localisée sur les filaments d'actine (5%),
- la troponine, localisée sur ces mêmes filaments (3%),
- l'x-actinine, qu'on retrouve au niveau des bandes Z (7%),
- la B-actinine, dans les filaments d'actine (1%),
- la protéïne M, au niveau de la ligne M du sarcomère (< 1%)
- la protéïne C dont le rôle serait de maintenir l'intégrité du sarcomère.
2.1) Le sarcomère.
Lorsque l'on observe des fibres musculaires au microscope, celles-ci
présentent une striation transversale caractéristique. C'est
l'alternance de bandes claires et de bandes sombres qui donne cet
aspect strié au muscle squelettique. La région claire est appelée
bande I ; I comme isotrope (qui laisse passer la lumière) et la région
sombre est appelée bande A ; A comme anisotrope ( qui ne laisse pas
passer la lumière). La ligne Z de l'allemand swishen signifie "entre",
coupe la bande I en deux et rejoint le sarcolemme pour donner de la
stabilité à toute la structure. L'unité répétitive comprise entre deux
lignes Z constitue le sarcomère, l'unité fonctionnelle de la cellule
musculaire. Les filaments d'actine et de myosine sont principalement
impliqués dans le processus de la contraction musculaire. Les
filaments d'actine et de myosine se chevauchent au niveau de la
bande A dont le centre s'appelle zone H à cause de sa plus faible
densité optique due à l'absence de filaments d'actine dans cette

J-P. Physiologie du muscle.
-4-
région. La zone H est divisée en deux par la ligne M qui indique de ce
fait le centre du sarcomère. La ligne M est formée de la superposition
des structures protéïques qui soutiennent les filaments de myosine.
méromyosine légère lourde
molécule de méromyosine
troponine
tropomyosine
molécules d'actine
FIBRE MUSCULAIRE
myofibrille
bande I bande A
ligne Z ligne Z
ligne Z ligne Z
SARCOMÈRE
MYOFIBRILLE
zone H
ligne M

J-P. Physiologie du muscle.
-5-
2.1.1) Caractéristique des myofilaments.
Grâce à leur propriété chimique et physique, les filaments de
myosine et d'actine sont particulièrement impliqués dans les
processus de la contraction musculaire.
La myosine est une molécule beaucoup plus grosse que l'actine
comportant des têtes globulaires qui saillent sur les côtés pour
former les ponts transversaux. Cette partie du filament de
myosine porte le nom de méromyosine lourde. Il existe environ
200 molécules de méromyosine pour un filament épais de
myosine. Chaque tête globulaire contient un site de liaison
capable de se lier à un site d'une molécule d'actine. Les
molécules de myosine des deux moitiés de chaque filament
épais sont orientées dans des directions opposées, de telle
sorte que toutes les extrémités de leurs tiges sont dirigées vers
le centre de la fibre. Grâce à cette disposition, les effets
moteurs des ponts aux deux extrémités de chaque filament
épais sont dirigés vers le centre, déplaçant ainsi les filaments
minces aux deux extrémités du sarcomère vers le centre du
sarcomère au cours du raccourcissement.
La tête globulaire de la myosine contient aussi un site
enzymatique séparé qui catalyse la dégradation de l'ATP en
ADP et en phosphate inorganique, libérant l'énergie chimique
mise en réserve dans l'ATP. La scission de l'ATP se produit sur
la molécule de myosine avant qu'elle se fixe à l'actine, mais
l'ADP et le phosphate inorganique produits demeurent liés à la
myosine.
molécule de myosine pont site de liaison
de l'actine
site ATPase
de la myosine
Filament de myosine
En conclusion la myosine peut grâce à ses ponts transversaux
entrer en combinaison chimique avec l'actine et agir comme
enzyme pour hydrolyser l'ATP. Cette réaction s'accompagne de
libération d'énergie, laquelle est pour une part utilisée en vue
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
6
7
8
9
10
11
12
13
14
15
16
17
18
19
1
/
19
100%