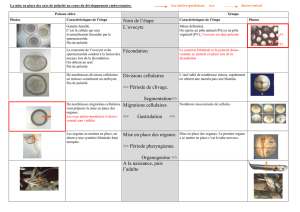
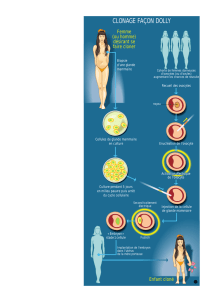
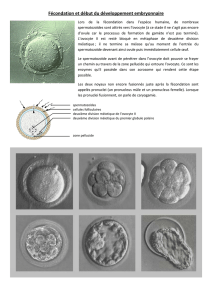
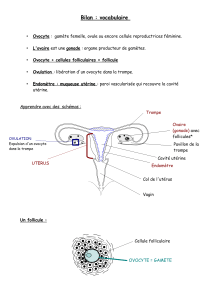
Déterminisme dorsa-ventrale de drosophile de la polarité de l`embryon

S SYNTHÈSE
1992 8
Eliane Mohier
directeur recherche au Cnrs.
cac
t,
dorsal. dosai
dorsal,
dorsal.
°

L

Y,
J
1
à
à
à

/
°
1
I
(1)

 6
7
8
9
10
6
7
8
9
10
1
/
10
100%