l`embryologie devenue moléculaire - iPubli

6
ÉDORL
Franço Jacob
RÉFÉRENCES·-
·
ADRESSE
L
»

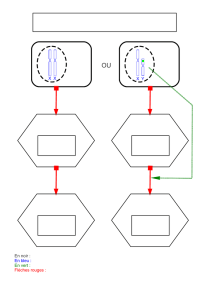
in situ
• 7
1
/
2
100%